

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Это наиболее гетерогенная из всех 7 групп. Хромосомы 4А (IV) и 4Д (ХV) обладают сходным действием, хромосома 4В (VIII) резко отличается от них. Нуллисомики по всем трем хромосомам имеют узкие листья и нежные стебли. Зрелые растения карликовые и мужские стерильные.
Гомеологичная группа 5 (хромосомы: 5В (V), 5A (IХ), 5Д (ХХIII))
Хромосомы этой группы имеют общую тенденцию увеличивать диаметр стебля и ширину листьев. Все нуллисомики имеют узкие листья и тонкие стебли, у тетрасомиков стебли толще, чем у нормальных растений. Все нуллисомики позднеспелые. Их колосья уменьшенного размера и имеют мелкие чешуи и зерно. Они фертильны по женской линии и стерильны по муской. Хромосома 5Д оказывает наиболее сильное влияние, хромосома 5А немного более сильное, чем хромосома 5В.
Гомеологичная группа 6 (хромосомы: 6А (Х), 6A (VI), 6Д (ХIХ))
Нуллисомики этой группы характеризуются узкими листьями, нежными стеблями и узкими, выступающими колосковыми чешуями, придающими колосьям странный вид. Все три нуллисомика женские фертильны, хотя фертильность может снижаться вследствие пистиллоидности. Нулли 6А и особенно, нулли 6Д до некоторой степени мужски фертильны. Моносомики, трисомики и тетрасомики мало отличаются от нормальных. Хромосома 6Д наименее существенна в этой группе, хромосома 6В возможно наиболее существенна. Нуллисомики по этой хромосоме у безостого сорта Чайниз Спринг имеют длинные ости.
Гомеологичная группа 7 (хромосомы: 7В (VII), 7A (ХI), 7Д (ХХI))
Нуллисомики этой группы мало отличаются от нормальных растений в начале развития. Зрелые различимы по незначительному уменьшению мощности развития и высоты и некоторым особенностям колос. Фертильность нулли 7А и 7Д почти нормальная, у 7В уменьшенная в связи с пистиллоидностью. Моносомики и трисомики в основном нормальные, все тетрасомики имеют уменьшенные размеры и фертильность. Они относятся к самым слабым по развитию из всех серий.
Нуллисомики по всем группам являются карликами и полукарликами: их высота колеблется от 1/3 до 1/5 высоты нормальных растений, листья их короче и уже листьев нормальных растений за исключением нуллисомиков по хромосоме 2А (2 группа), растения которой имеют короткие, но широкие листья. Нуллисомики 2 группы фертильны по мужской линии, но стерильны по женской, полностью стерильны в группе 1, мужски стерильны по группам 4 и 5, фертильность заметно снижена по сравнению с нормальными растениями в группах 6 и 7. В группах 6 и 7 фертильность снижется в связи с пистиллоидностью. Нуллисомики по группам 4,5,6 имеют тонкую, нежную соломинку.
Интересно отметить, что нуллисомики Чайниз Спринг по хромосомам 4В и 6В имеют длинные прямые ости. Отмечено, что у нуллисомиков по хромосоме 4А (4 группа) 4Д пыльники растрескиваются, пыльца имеет нормальный вид, но не способна к оплодотворению.
Тетрасомики часто имеют пониженную фертильность. Особенно это проявляется у тетрасомиков по хромосомам 4а, 7А, 1В и 2В. В условиях, благоприятных для развития растений, большинство моносомиков сорта Чайниз Спринг не отличаются от дисомиков, но при менее благоприятных обстоятельствах они отклоняются от нормальных в сторону соответствующих им нуллисомиков. У одних сортов пшеницы фенотипы моносомиков могут более отклоняться от нормы, чем у других.
Из всего набора моносомных линий Чайниз Спринг только две, а именно 5А и 5Д, фенотипически отличаются от дисомиков.
8 лекция
Методы создания серий моносомных линий – 2 часа
Использование анеуплоидов для генетических исследований пшеницы проводится в основном по трем направлениям: 1) создание новых серий моносомных линий у перспективных сортов; 2) моносомный анализ качественных и количественных признаков; 3) замещение хромосом.
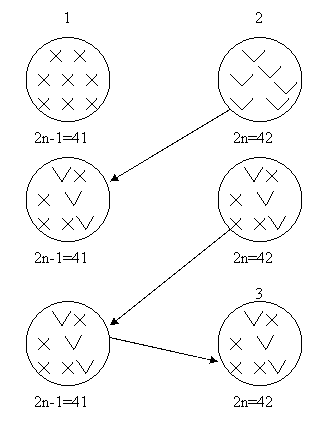
Новые серии моносомных линий создаются для сортов, наиболее приспособленных к определенным природным условиям, а также для тех, по которым в дальнейшем предполагается создание замещенных линий. Чаще всего это делают на основе набора анеуплоидов сорта Чайниз Спринг. В современной практике моносомные линии любого сорта получают путем возвратных скрещиваний с каждым из моносомиков. Работа ведется по следующей схеме:
1.Растения Чайниз Спринг, моносомные по каждой из 21 хромосомы, скрещиваются с сортом, у которого создается новая серия. Моносомные растения используются в качестве материнской формы. В потомстве появляются растения двух типов – моносомные и дисомные по данной хромосоме.
2.Отобранные путем цитологического анализа растения все линий скрещивают вновь с тем же сортом, т. е. осуществляются первое возвратное скрещивание, или первый беккросс. Потомство от него получается опять смешанное, состоящее из моносомиков и дисомиков.
3.Моносомные растения в каждом поколении скрещивают возвратно с тем же сортом не менее шести раз до воссоздания генотипа скрещиваемого сорта. Полученный в шестом беккроссе моносомик генетически почти идентичен отцовскому сорту. Унивалентная хромосома, как правило, остается идентичной материнскому сорту. Представленный ниже рисунок поясняет создания серии моносомных линий (рис. 2):
 | |
| |
Работа с анеуплоидами пшеницы требует постоянного контроля количества хромосом в клетках каждого растения, так как моносомные растения при благоприятных условиях выращивания не отличаются о дисомных по генотипу. Исключение составляют растения Чайниз Спринг по хромосоме 5А, которые имеют спельтоидный колос, на что указывали многие исследователи. Моносомные растения по хромосоме 5Д при обычном весеннем посеве отличаются от нормальных позднеспелостью. В новой серии моносомных линий по сорту Казахстанская 126 выделены 6 хромосом: 3А, 5А, 7А, 4В, 6В и 5Д, каждая из которых несет отдельно маркерные гены, определяющие треугольный вырез у основания ости, спельтоидный тип колоса, узкоскошенное скошенное плечо, остистость у гибридов F1, позднеспелость соответственно, что также дает возможность выделять моносомики по фенотипу без цитологического анализа. Несмотря на то, что иногда удается выделить моносомные растения без цитологического анализа, исследования с использованием анеуплоидов требует постоянной и кропотливой работы по цитологическому контролю количества хромосом.
9 лекция
Возникновение линии с телоцентрическими и изохромосомами – 2 часа
Другой вид цитологических нарушений, обнаруженных Сирсом в кариотипе пшеницы Чайниз Спринг привел к созданию телоцентрической хромосомы. Неправильные поперечные деления хромосом – унивалентов в районе центромеры как в первом, так и во втором делениях мейоза приводят к утере одного из плечей хромосомы и образованию телоцентрической хромосомы. Телоцентрические хромосомы часто образуют затем изохромосомы. Последние формируются при редупликации телоцентрических хромосом и развертывании образовавшихся при удвоении хроматид на 1800 (рис. 1). Телоцентрические хромосомы можно идентифицировать в М1 по их малому размеру и отсутствие изгиба, но легче всего телоцентрики иденифицировать в А1, когда они располагаются между двумя полюсами под прямым углом к оси веретена, с центромерой в терминальном положении.
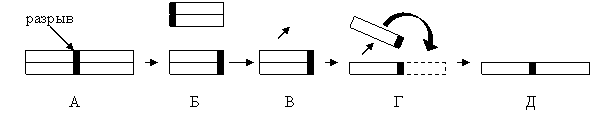
Рис. 1. Вероятное происхождение изохромосом из унивалентов. А - исходоне состояние и разрыв хромосомы; Б и В – телоцентрические хромосомы; Г и Д – возникновение изохромосомы
Изохромосому значительно легче отметить в М1, чем в А1. Обычно, она лежит вне пластинки и бывает толще, чем цельная моносома, форма ее варьирует от круглой до овальной в зависимости от длины плеча, по которому она образовалась.
Влияние 5В хромосомы мягкой пшеницы на конъюгацию хромосом - 2 часа
Выше было показано, что гомеологичные хромосомы, происходящие от разных диплоидных предков мягкой пшеницы генетически родственны. Однако в мейозе конъюгации между гомеологичными хромосомами разных геномов не наблюдается, а всегда образуется 21 бивалент в результате конъюгации только гомологичных хромосом. Отсутствие конъюгации между гомеологичными хромосомами объясняется подавлением процесса конъюгации между ними активностью 5В хромосомы.
Сирс и Окамото (1958), Райли и Чепман (1958) было установлено, что в присутствии 5В хромосомы у мягкой пшеницы в конъюгацию вступает только гомологи, а в отсутствие ее образуются мультиваленты. Ими также было установлено, что механизм управляющий образованием только бивалентов, эффективен и в гемизиготном состоянии. Поскольку изучение всех остальных нуллисомиков не обнаружило влияние на мейоз, сходного с действием 5В хромосомы, был сделан вывод, что ген, подавляющий конъюгацию гомеологичных хромосом, не дублирован ни водной другой хромосоме T. aestivum. Знание этой системы позволяет в какой-то мере использовать ее в практической селекции. Так отсутствие 5В хромосомы пшеницы при межродовых скрещиваниях может облегчить передачу генов, ответственных за хозяйственно-ценные признаки от близких родов мягкой пшеницы. Следовательно, генетические разнообразие ржи, эгилопса и пырея может быть использовано для улучшения пшеницы.
10 лекция
Использование серий нуллисомиков и моносомиков для генетического анализа – 4 часа
Для создания серии анеуплоидов изучение количественных признаков связанных с продуктивностью растений, осуществлялось исключительно путем гибридологического анализа. Однако, изучение генетики количественных признаков традиционным методом гибридологического анализа не дает возможности локализовать гены, контролирующие эти признаки в определенных хромосомах.
При проведении моносомного анализа, сорт по которому предстоит выявить генетический эффект отдельных хромосом в развитии определенного признака, скрещивают с каждой из 21 моносомных линий используемых в качестве женского родителя. Из популяции дисомиков и моносомиков в поколении F1 цитологически идентифицируются и выделяются момносмные растения, у которых унивалентная хромосома происходит от исследуемого сорта – донора. Если моносомный родитель гомозиготен по рецессивному аллелю и рецессивный признак проявится в определенной линии в гемизиготном состоянии (моносомном), то анализ моносомных растений в поколении F1 позволяет локализовать рецессивный ген, контролирующий исследуемый признак.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
