

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
После заключительного беккросса моносомики самоопыляют и в последующем поколении отбирают дисомные растения (примерно 24%), у которых в соматических клетках содержится 40 хромосом от Чайниз Спринг и одна пара хромосом от сорта – донора. Для получения полной серии замещения подобная работа должна проводиться по каждому из 21 нуллисомиков.
Описанный метод 1 имеет ограниченное применение, так как большинство нуллисомиков, за исключением 7В,7Д, 1В представляют собой растения стерильные, что затрудняет их свободное использование в скрещиваниях.
Метод 2
1этап. В каждой из моносомных линий реципентного сорта отбирают моносомное растение и опыляют его донорским сортом. Из смешанного потомства F1 путем цитологического анализа выделяют моносомики.
2этап. Проводится самоопыление моносомиков F1, отбирают дисомики, которые гомозиготны по хромосоме донора.
3этап. Применяют возвратное скрещивание моносомиков каждой линии с гомозиготным по донорской хромосоме дисомиком F2. Соответствующий моносомик при этом представляет собой «повторного родителя».
4 этап. Потомство от беккроссов снова изучают цитологически, отбирают моносомики и их дисомное потомство, получаемое от самоопыления, скрещивают с соответствующим моносомиком реципентного сорта, который используют как материнскую форму. Эти процессы самоопыления и скрещивания повторяют до тех пор, пока генотип реципента по 20 хромосомам не восстанавливается на 98-99% через 12 чередующихся поколений беккросса самоопыления: унивалентная хромосома от сорта донора остается неизменной. Ниже представлена схема замещения хромосом:
Р: (♀) 20II К.126 + IК.126 х ♂21II тимштейн
дисомик моносомик
20К.126 + 1 К.126 20 К.126+1 тимштейн
20 тимштейн 1 тимштейн 20тимштейн
ВСI 20II К.126 + I К.126 х К.126 + I тимштейн
20 тимштейн
Последний тип скрещивания повторяют в течение пяти-шести поколений. На заключительном этапе отбираю моносомики, после их самоопыления выделяют в потомстве дисомики 20II К.126 + III тимштейн.
Метод 3
Реципентной формой является серия монотелоцентрических линий.
Схематично работу по методу 3, когда реципентном избрана серия монотелоцентриков Чайниз Спринг (Ч. С.), а сортом – донором Казахстанская 126 (Каз.126), можно изобразить следующим образом:
Р: ♀ 20II Ч. С. + Тело – Ч. С. х ♂ 21II К.126
дисомик моносомик
20 Ч. С. + 20 Ч.С. + I К.126
20 К.126 I К.126 20 К.126
ВСI 20II Ч. С. + х 20 Ч. С. + I К.126
20 К.126
Последний тип скрещивания повторяют в течение 5-6 поколений. В заключительном этапе отбирают моносомики, после их самоопыления выделяют в потомстве дисомики 20II Ч. С. + I К.126.
12 лекция
Замещение хромосомы, вовлеченной в реципрокную транслокацию – 2 часа
Влияние транслокации на процесс замещения хромосом показано на примере скрещивания моносомика Чайниз Спринг по хромосоме Х (♀) с сортом Тэтчер (♂). У моносомных растений F1 в метафазе I образовалось 19 бивалентови цепь из трех хромосом IV о Чайниз Спинг, IV-X и X-IV от Тэтчер.
Если хромосомы в метафазе I образуют цепь из трех хромосом, то в результате их расхождения в анафазе 20- хромосомные гаметы гибрида будут нести хромосому IV от Чайниз Спринг, а 21 – хромосомные гаметы – хромосомы с транслоцированными сегментами IV-X и X-IV от сорта Тэтчер. Поскольку при самоопылении гибридов F1 эти 21 хромосомные гаметы образуют потомство дисомики, используемые для беккроссирования с моно - Х Чайниз Спринг, моносомные растения в F1 и последующих поколениях беккросса будут иметь следующую хромосомную конституцию: 19II + цепь их трех хромосом. В участках хромосом IV сорта Чайниз Спринг, будет происходить кроссинговер в F1 и во всех поколениях беккроссах и гены, полученные от сорта донора будут замещаться аллелями от Чайниз Спринг. Замещающаяся хромосома от Тэтчер, гомологичная паре хромосом Х сорта Чайниз Спринг, будет делиться между двумя парами хромосом IV-X и X-IV. Схему замещения хромосом можно изобразить следующим образом:
РI моно-Х Чайниз Спринг (♀) х Тэтчер (♂)
F1 19 Ч. С. + Ч. С. IV + Т. Х - IV
19 Т. Т. IV-X
В F1 и последующих беккроссах с моно-Х чайниз Спринг (♀) при опылении наиболее вероятно функционирют следующие типы гамет: IV-X + X-IV; Ч. С. IV + X-IV; Ч. С. IV + IV - X. В потомстве беккросса образуются моносомики трех типов:
19 Ч. С. + Ч. С. IV + Х - IV; 19 Ч. С. + Ч. С. IV + Х - IV;
19 Т. Т. IV-X 19 Ч. С.IV
19 + Ч. С. IV + IV - Х.
19 Ч. С.IV
У каждого из моносомиков образуется в метафазе I цепь из трех хромосом. Частота кроссинговера будет зависеть от степени сходства транслоцированных сегментов с соответствующими участками гомологичных хромосом анеуплоидной пшеницы.
13 лекция
Метод реципрокного замещения хромосом – 2 часа
Возможность создания моносомных линий с одновременным получением серий с теплоцентрическими хромосомами и реципрокным замещением обсуждается в работах Ло (1968). Автор предлагает при создании моносомных линий скрещивать изучаемый сорт с моносомиками Чайниз Спинг, а затем использовать моносомное растение F1 не только для возвратного скрещивания с исходным сором, но и одновременно скрещивать момносомики из F1 с дителоцентриком Чайниз Спринг по хромосоме, переводимой в моносомное состояние. Ниже приведена схема одновременного создания моносомной и монотелоцентрической линий:
В результате получаемые монотелоцентрические растения позволяют одновременно проверять отобранный в F1 моносомик для выявления «смены унивалента» и начать работы по замещению гемизиготной линией Чайниз Спринг. Замещение может проводиться вместе с беккрссированием для получения моносомной серии, н будет задерживаться на год, т. е. отставать на одно поколение. В результате можно создать полную серию моносомных линий у сорта А и на одно поколение позже – 21 линию монотелоцентриков. Эта методика сокращает время на создание реципрокного замещения хромосом. Возможность проведения такой схемы замещения будет зависеть от того: 1) будет ли у других сортов жизнеспособны растения, у которых утеряны плечи хромосом и замещены телоцентрической хромосомой от Чайниз Спринг;
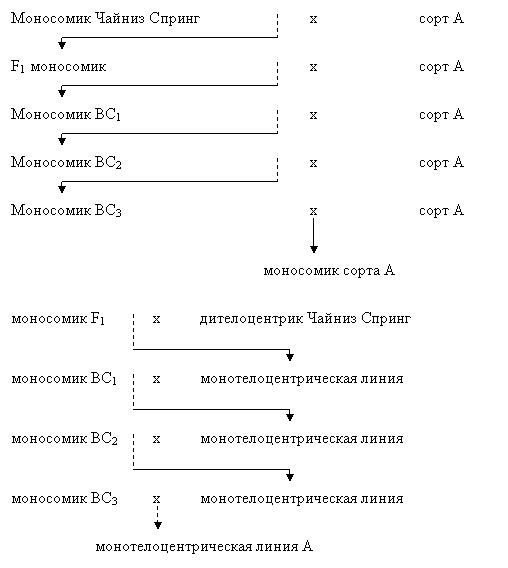
2) будут ли монотелоцентрические линии достаточно фертильными при использовании их пыльцы для опыления.
14 лекция
Метод создания серий с межсортовым замещением хромосом – 2 часа
После получения набора моносомных линий желаемого сорта хромосомы от других сортов могут быть перенесены путем создания линий с замещением хромосом. Будут рассмотрены 3 метода получения замещенных линий, которые обсуждались в широко известных работах Сирса и Унрау с сотр. Эти методы различаются в основном типом анеуплоида, который используется в качестве материнской родительской формы для повторных скрещиваний. Материнскую или отцовскую форму, используемую многократно при возвратных насыщающих скрещиваниях, называют «Повторным родителем». В первом методе таким «повторным родителем» является нуллисомик, во втором методе – моносомик (унивалент при этом – целая двуплечая хромосома), в третьем методе – монотелоцентрик (унивалент - телоцентрическая, одноплечая хромосома). Основная схема замещения хромосом одинакова для всех трех методов.
Метод 1.
1 этап. Нуллисомик Чайниз Спринг, у которого отсутствует определенная пара хромосом, скрещивается в качестве материнского компонента с избранным сором донором.
2 этап. Все растения F1 – моносрмики, причем унивалентная хромосома происходит от сорта донора. Моносомики F1 по данной хромосоме скрещивают как отцовский компонент с нуллисомиком (♀) Чайниз Спринг («повторный родитель»).
3 этап. В пототстве ВСI от первого возвратного насыщающего скрещивания с «повторным родителем» имеются два типа растений: примерно 94-99% моносомиков с унивалентной хромосомой от сорта –донора и 1-6% нуллисомиков. Пыльцой моносомных растений снова опыляют соответствующие нуллисомики Чайниз Спринг. Подобные насыщающие скрещивания проводят шесть-восемь раз в зависимости от того, насколько сильно различают меду собой генотипы сортов донора и реципента.
После заключительного беккросса моносомики самоопыляют и в последующем поколении отбирают дисомные растения (примерно 24%), у которых в соматических клетках содержится 40 хромосом от Чайниз Спринг и одна пара хромосом от сорта – донора. Для получения полной серии замещения подобная работа должна проводиться по каждому из 21 нуллисомиков.
Описанный метод 1 имеет ограниченное применение, так как большинство нуллисомиков, за исключением 7В,7Д, 1В представляют собой растения стерильные, что затрудняет их свободное использование в скрещиваниях.
Метод 2
1этап. В каждой из моносомных линий реципентного сорта отбирают моносомное растение и опыляют его донорским сортом. Из смешанного потомства F1 путем цитологического анализа выделяют моносомики.
2этап. Проводится самоопыление моносомиков F1, отбирают дисомики, которые гомозиготны по хромосоме донора.
3этап. Применяют возвратное скрещивание моносомиков каждой линии с гомозиготным по донорской хромосоме дисомиком F2. Соответствующий моносомик при этом представляет собой «повторного родителя».
4 этап. Потомство от беккроссов снова изучают цитологически, отбирают моносомики и их дисомное потомство, получаемое от самоопыления, скрещивают с соответствующим моносомиком реципентного сорта, который используют как материнскую форму. Эти процессы самоопыления и скрещивания повторяют до тех пор, пока генотип реципента по 20 хромосомам не восстанавливается на 98-99% через 12 чередующихся поколений беккросса самоопыления: унивалентная хромосома от сорта донора остается неизменной. Ниже представлена схема замещения хромосом:
Р: (♀) 20II К.126 + IК.126 х ♂21II тимштейн
дисомик моносомик
20К.126 + 1 К.126 20 К.126+1 тимштейн

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
