
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При попадании в желудок мамонта 30-40 литров холодной воды с температурой 2-3 градуса, необходимо еще дополнительное количество тепловой энергии для ее нагрева до температуры, при которой она может включаться в физиологические реакции организма. Кроме того, снижение температурной среды в желудке мамонта, на какой-то период могло вызывать замедление активности беспозвоночных симбионтов мамонта. Если при падении температуры в желудке мамонта происходила гибель части симбионтной флоры, то это могло очень негативно сказаться на усваивании пищи и могло вызвать ослабление животного.
Существенным ограничением является отсутствие в снеге минеральных веществ. Необходимость в минеральных веществах достаточно высока у всех крупных растительноядных групп млекопитающих. У гигантских млекопитающих с массой тела более 1 тонны зависимость от минеральных веществ еще выше. Внесезонная литофагия хорошо известна у современных африканских слонов (Shoshani et al., 1991; Di Silvestro, 1991). Усваивание питательных веществ растительноядными животными образующихся при расщеплении клетчатки и само ее расщепление происходит при химических реакциях на мембранах клеток. Эти физиологически реакции невозможны без калия, кальция, натрия, других микроэлементов, которые растительноядные млекопитающие получают из воды, или поедая минеральные соединения на солонцах (Паничев, 1990). В Евразии литофагия у M. primigenius была установлена на материале из местонахождений бассейна р. Колымы (Шило и др., 1983). Поедание глинистых минералов установлено также по содержимому желудков взрослых мамонтов (Шадринского и Юрибейского), киргиляхского детеныша мамонта и второго Ямальского детеныша (Люба, ЯМН № 000) (Верещагин, 1981; Верещагин, Николаев, 1982; Юрибейский мамонт, 1982; Косинцев и др., 2010).
Отсутствие не замерзающей воды и восполнение ее снегом у северного оленя (Rangifer tarandus Linnaeus 1773), в сочетании с невозможностью найти под глубоким снегом необходимые минеральные вещества, вызывает сезонное зимнее вымывание калия и кальция из костей скелета и частичное разрыхление костной ткани (Барабаш-Никифоров, Формозов, 1963). Этот сезонный минеральный дефицит преодолевается северным оленем с наступлением зимы и у особей, не имеющих других заболеваний полностью компенсируется в течение лета и не выходят за рамки физиологической нормы реакции у этого вида. У шерстистого мамонта сезонные изменения плотности костей скелета также наблюдаются, но неправильно интерпретируются как различные формы заболеваний опорно-двигательной системы (Лещинский, 2009).
Наиболее вероятным является гипотеза о том, что поедание снега зимой не было для шерстистого мамонта основным источником получения воды. Данные со стоянок средней и поздней поры верхнего палеолита на Русской Равнине говорят о том, что в местах скопления костей мамонтов существовали не замерзающие зимой источники воды, которые использовались, как шерстистым мамонтом, так и другими крупными млекопитающими. Как правило, такие родники расположены в основании бортов долин и участках разломов, что и было установлено на стоянке Костенки, а так же предполагается для некоторых других стоянок (Sinitsyn, Hoffecker, 2006; Anikovich, et al., 2007; Holliday, et al., 2007; Мащенко, 2009).
Приведенные примеры, поедание снега у современных растительноядных копытных вынужденная форма поведения, тяжело сказывающаяся на физиологии, но компенсируются за счет ширины нормы физиологических реакций у крупных млекопитающих. Полностью исключать такое поведение для шерстистого мамонта нельзя, скорее всего, мамонт зимой старался поедать наиболее сочные подснежные корма, хотя, данных об этом нет. Кроме того говорить о массе сочного подснежного корма для позднего плейстоцена Северо-востока Азии не приходится, учитывая либо полное отсутствие снега, либо его глубина не превышала 15-20 см.
Строение и форма расширения хобота у шерстистого мамонта - морфологический признак, отличающий вид M. primigenius, от современных слонов. В данной работе рассматриваются три случая наличия указанной морфологической особенности строения хобота у трех разных особей, разного индивидуального возраста (детеныш, молодая половозрелая особь и старая половозрелая особь). Это говорит о том, что представлена не случайная индивидуальная особенность, а признак, отмечаемый на разных стадиях онтогенеза. Данную черту морфологии хобота можно без сомнения рассматривать, как одну из адаптаций к жизни в условиях холодного климата. Вместе с тем, достаточно обоснованная и всеобъемлющая интерпретация функционального значения этой особенности морфологии хобота M. primigenius возможно, еще предстоит в будущем.
Мамонты с местонахождения Мус Хая
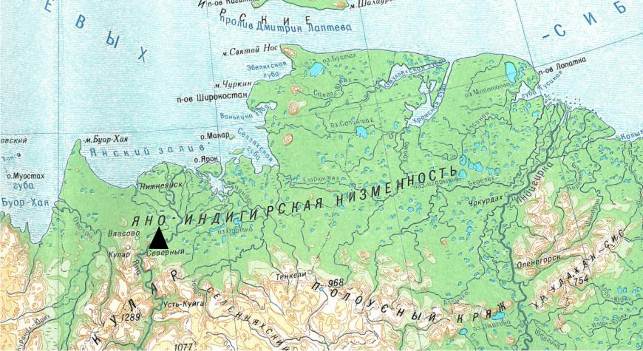
Устье безымянной реки, где был найден исследуемый костный материал, находится недалеко от местонахождения Мус Хая, левый берег р. Яна, нижнее течение. Мус Хая расположена в 30 км ниже по течению от пос. Северный, Усть-Янского улуса. Участок Мус Хая представляет собой обнажения жильных льдов протяженностью 1300-1400 м, с постоянно смываемым берегом. Высота от уровня воды составляет 60-70 м. Исследуемое местонахождение расположено на изгибе реки чуть выше места разделения реки на два русла (рис. 27.) (70°43' с. ш. 135°24' в. д.). Здесь в 2008 году были обнаружено большое скопление костей разновозрастных особей вида M. primigenius.
|
Рис. 27. Карта расположения местонахождения Мус Хая |
В 2008 году на местонахождении работала группа ученых из Геологического института РАН, института исследования Арктики и Антарктики и института материальной исторической культуры РАН. По их данным здесь было собрано около 1032 единиц костного материала вида M. primigenius, которые принадлежали примерно 26 особям. Кроме того, на местонахождение было найдено не большое количество остатков других представителей мамонтовой фауны: B. priscus; E. lenesis; C. antiquitatis; R. tarandus; U. arctos (Basilyan et al., 2011).
В 2010 году на местонахождении Мус Хая нами исследовано 52 единицы костных остатков M. primigenius. Мы полагаем, что это далеко не последние находки данного местонахождения, т. к. по мере размыва берегов р. Яна с каждым годом вскрываются новые «залежи» остатков древней фауны.
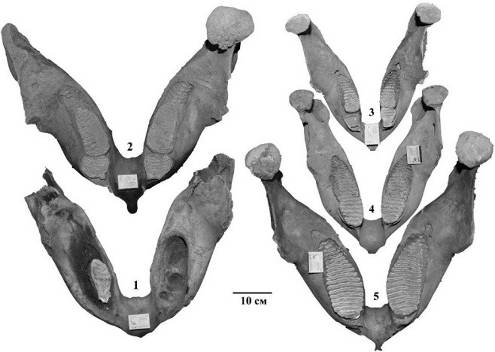
На местонахождении были найдены в основном остатки M. primigenius и в небольших количествах C. antiquitatis и B. priscus. Остатки мамонтов принадлежат особям разного биологического возраста. Об этом свидетельствуют размеры костей конечностей, степень срастания проксимальных и дистальных эпифизов, а также срастание краниальных и каудальных эпифизов и апофизов позвонков. Кроме того, на местонахождении были обнаружены несколько нижних челюстей мамонтов с генерацией зубов dp3-dp4, dp4-m1, m2-m3 и m3. Нужно также отметить челюсть очень пожилой особи №СГ-1 (рис. 28.1), у которой на челюсти сохранился почти полностью стертый правый коренной зуб (m3/dex), количество пластин 4 шт., толщина эмали 2,1 мм (по направлению к корням толщина эмали увеличивается). Левый зуб, видимо выпал при жизни. Такой вывод сделан, исходя из того, что альвеола не имеет видимых механических повреждений, которые обычно наблюдаются у экземпляров с посмертным выпадением зубов. Остаток правого коренного зуба расположен посередине альвеолы, а пространство между краями альвеолы и корнем зуба затянуто костной тканью. Аналогичная находка была сделана на Севском местонахождении (Брянская область) (Мащенко, 1992), где индивидуальный возраст особи был определен как более 60 лет. Таким образом, индивидуальный возраст экз. №СГ-1 видимо, составляет около 60-70 лет, т. е. глубокая старость. На нижней челюсти №СГ-2 сохранились зубы генерации m2 и m3 на обеих ветвях. Зубы m3 находятся на стадии прорезания, стиранием затронуты 13 пластин на правом и 14 на левом (рис. 28.2). На обоих m3 в видимой части имеется по 20 пластин, остальные пластины еще не прорезались. Зуб генерации m2 почти полностью стерт. На правом зубе сохранились пять пластин на левом шесть. Мезиальная часть коронки стерта до основания (поверхность гладкая). Частота пластин на 10 см - 10. На экз. №СГ-3 сохранились зубы второй и третьей генерации (dp3-dp4) (рис. 28.3). Зубы dp4 находятся на стадии прорезания. На левом dp4 имеется 14 пластин на правом 13. Стиранием затронуты 8 пластин на левом и 9 пластин на правом. Зубы генерации dp3 находятся на стадии полного стирания, правый зуб выпал. На зубе сохранились четыре пластинка. Мезиальная часть зуба полностью стерта и на нем образована гладкая поверхность. На экз. №СГ-4 (рис. 28.4) сохранились зубы третьей и четвертой генерации (dp4-M1). На зубах третьей генерации (dp4) сохранились по 6-7 пластин. Зубы четвертой генерации (M1) находятся на стадии прорезания. На обоих M1 имеется по 14 пластин из них стиранием затронуты 8 пластин на левом зубе и 8 на правом. Индивидуальный возраст этой особи от 14 до 17 лет. Экз. №СГ-5 имеет превосходную сохранность (рис. 28.5). На челюсти сохранились зубы шестой, последней генерации – m3. Частота пластин на 10 см - 9. На обоих зубах по 20 пластин, остальные пластины скрыты в челюсти. Стиранием затронуты 17 пластин на обоих зубах. Индивидуальный возраст от 20-21 до 27 лет.
|
Рис. 29. Выборка нижних челюстей (mandibula) мамонтов обнаруженные на местонахождении Мус Хая: 1. - №СГ-1; 2. - №СГ-2; 3. - №СГ-3; 4. - №СГ-4; 5. - №СГ-5. |
Первый шейный позвонок мамонта (№СГ-8) темно-коричневого цвета, местами имеются желтоватые оттенки, апофизы облитерированы. Экз. №СГ-9 первый шейный позвонок коричневого цвета, хорошей сохранности, апофизы облитерированы. Первый шейный позвонок №СГ-10, цвет темно-коричневый, области реберно-поперечных отростков обломаны. Экз. №СГ-11 коричневого, желтоватого цвета. Правый апофиз позвонка отсутствует в связи с тем, что был не приросший. Промеры позвонков представлены в таблице 18. Сравнение исследуемых экземпляров с таковыми из других местонахождений показывает, что индивидуальный возраст этих особей может составлять не менее 18-19 и не более 27-30 лет.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
