
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Накапливание жира в подкожной области шеи и передней части холки впервые было установлено при изучении оймяконского детеныша (АН № 000) (Боескоров и др. 2007). Эта особенность физиологии М. primigenius принципиальна для выживания детенышей первого года жизни зимой при недостатке пищи. Другой особенностью ранних стадий постнатального онтогенеза шерстистого мамонта является ускоренная смена зубов первых смен (DP2, DP3) по сравнению с современными слонами (Кузьмина, Мащенко, 1999; Maschenko, 2002). Быстрое начало стирания зубов третьей смены (DP4) в возрасте около 2-2,5 лет и начало стирания зуба второй смены (DP3) в возрасте 4-6 месяцев указывают на начало перехода к питанию кормом взрослых особей. Эта особенность шерстистого мамонта не может быть объяснена продолжительным периодом лактации с продолжительностью как у современных слонов. Обе указанные особенности показывают о более коротком периоде активной лактации у М. primigenius, который продолжался до наступления зимы, когда детеныши достигали возраста 6-8 мес. В зимний период уровень лактации вероятно, еще больше снижался. В противоположность М. primigenius у L. africana уровень лактации не снижается в течение первого года и лактация продолжается больше 2 лет (Shoshani et al., 1991).
Еще одной адаптацией шерстистого мамонта к холодному климату в первые 6-8 мес. жизни является ускоренный рост по сравнению с ростом в тот же период жизни у современных слонов (Maschenko, 2002). Возможно, наблюдаемые различия в размерах тела у детенышей мамонтов сходного возраста (установленного по смене зубов) определяются тем, что у одних особей возможность ускоренного роста реализовывалась, а у других – нет.
У второго ямальского детеныша (ЯНМ № 000) депонирование жира происходило не только в области холки, но и в подкожном слое на боках туловища. Последнее так же было отмечено и у взрослого самца березовского мамонта (Заленский, 1903).
С физиологической точки зрения, накопление жира важно при подготовке к зимнему сезону и недостатке пищи. Увеличение количеств подкожного жира в течение лета отмечено у современной якутской лошади (Габышев, 1957) и других крупных млекопитающих арктической зоны. Скорее всего, при окончании периода вегетации растений (сентябрь) лактация у самок мамонта, имеющих детенышей, значительно сокращалась, а к концу зимы прекращалась полностью. С этим связано более раннее начало стирания зубов первой и второй смен (DP2, DP3), установленное у детенышей мамонтов Русской равнины (Кузьмина, Мащенко, 1999). Именно холодный сезонный климат обусловливает более ранний и более быстрый переход к питанию только растительной пищей детенышей мамонта, по сравнению с детенышами современных слонов. Различия в сроках стирания DP2, DP3 у детенышей мамонтов и слонов составляют около 6 мес.
Накопление жира в подкожном слое туловища установлено у двух (оймяконский и второй ямальский) из четырех изученных и хорошо сохранившихся детенышей. По-видимому, накопление подкожного жира зависело от питания, нормального физиологического развития и роста детенышей в семейной группе, также как у современных слонов. Структура популяций мамонтов (семейная группа из самок с детенышами, самцовые группы, одиночные самцы) была установлена для М. primigenius в Севске и М. columbi Falconer, 1857 в Хот-Спрингс (Южная Дакота) (Maschenko, 2002). Длительная забота о потомстве у млекопитающих, объединенных в семейные группы - один из важных элементов выживания. Например, истощение киргиляхского детеныша позволило (1979, 1981) вполне обоснованно предположить, что этот детеныш погиб после гибели матери, отстав от семейной группы.
На возрастание роли самостоятельного питания у детенышей М. primigenius при переходе от летнего сезона к зимнему может косвенно свидетельствовать содержимое толстого кишечника второго ямальского детеныша (Косинцев и др., 2010). Оно состояло главным образом из растительных остатков, которые практически не деструктурированы. Кроме того, значительную часть заполнения кишечника составляет илистый субстрат серого цвета. Помимо того, что усваиваемость растительной пищи слонами около 40%, наличие хорошо сохранившихся растительных остатков в толстом кишечнике объясняется возрастанием значения самостоятельного питания этой особи. Индивидуальный возраст этого детеныша не менее 3 мес., DP2 практически не стерт, DP3 полностью сформирован. Минеральное заполнение кишечника второго ямальского детеныша может свидетельствовать о начале регулярного поедания определенных типов породы, необходимых для нормального переваривания растительной пищи. В частности поедание карбонатов и глин отмечено для современных азиатских и африканских слонов (Shoshani et al., 1991) и мамонтов (Maschenko et al., 2003; Maschenko et al., 2007; Maschenko, 2007). Глинистые массы присутствуют в кишечниках киргиляхского и второго ямальского детенышей.
Наличие растительных остатков в кишечнике второго ямальского детеныша может быть следствием поедания помета взрослых особей. У детенышей африканских слонов направленный сбор и поедание помета взрослых особей отмечены с возраста 1-1,5 года (возможно, раньше), когда происходит переход на смешанное питание (Shoshani et al., 1991). Считается, что с пометом взрослых особей поступает микрофлора, необходимая для переваривания растительной пищи.
Ускоренные темпы роста детенышей мамонта в течение первых 6 мес. жизни, по сравнению с ростом детенышей современных слонов, были установлены по скелетам детенышей мамонта из Севской популяции. Определено, что рост новорожденных детенышей мамонта из Севска 700-750 мм (Maschenko, 2002; Maschenko et al., 2006), что заметно меньше, чем у новорожденных детенышей L. africana (до 1000 мм) (Laws, 1966; Sikes, 1966). У М. primigenius высота тела в холке в 6-8 мес. (1150-1180) соответствует таковой у L. africana в 12-14 мес. К двум годам различия в скорости роста и размерах тела у африканского слона и шерстистого мамонта почти полностью нивелируются (Maschenko, 2002).
Размеры трупов детенышей мамонта подтверждают первоначально зарегистрированные на скелетах из Севска данные об их ускоренном росте в первые месяцы жизни. Рост новорожденного первого ямальского детеныша 690 мм (см. табл. 4). У второго ямальского детеныша за первые 2-3 месяца жизни отмечено увеличение высоты тела по сравнению с первым 270 мм (39% от высоты тела новорожденного первого ямальского). Высота тела киргиляхского детеныша за 7 месяцев увеличилась на 50% от высоты тела первого ямальского детеныша. При сравнении с высотой тела первого ямальского детеныша высота тела хромского за 4-5 месяцев увеличилась на 210 мм (30%).
Продольное растягивание и поперечный разрыв туловища хромского детеныша трудно поддается реконструкции. Данное обстоятельство не вызвано нападением крупных хищников. При разрывании тела жертвы львами или медведями на шкуре остаются множественные следы от клыков и когтей, которые отсутствовали на трупе хромского детеныша. Выедание мягких тканей на шее, холке и голове произведено песцами (A. lagopus) при вытаивании трупа из вечной мерзлоты, и следы погрызов характерны именно для этого вида хищников. Таким образом, причины деформации туловища хромского детеныша пока остаются неясными.
Определение конкретных причин смерти детенышей мамонта очень затруднено. Имеющаяся на данный момент информация позволяет считать, что для большинства сохранившихся в вечной мерзлоте захоронений детенышей причинами смерти были либо травмы, не совместимые с жизнью (оймяконский, первый ямальский и хромской), либо гибель в воде или болоте (киргиляхский, первый ямальский).
Мамонты с р. Селлях
Местонахождение на р. Селлях является типичным захоронением шерстистых мамонтов, характерным для северных районов Якутии. Кости залегают в одном горизонте. Разброс костей относительно невелик, примерно в радиусе 10-12 м. Глубина залегания остатков 5-6 м. Около 99% всех обнаруженных нами костных остатков принадлежат молодым особям M. primigenius.
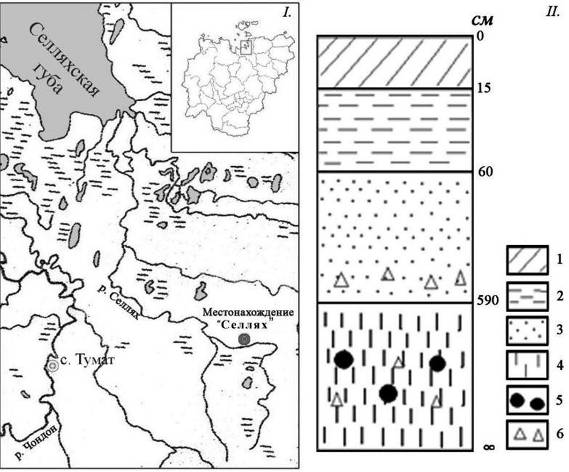
Местонахождение «Селлях» расположено на правом берегу р. Селлях (верхнее течение) в 57 км к востоку от п. Тумат Усть-Янского улуса (70°45 с. ш. и 140°45 в. д.) (рис.7.I).
Летом 2011 г. местные «недропользователи», занимающиеся сбором бивней мамонта, обнаружили кости древних животных, которые располагались на небольшом участке. На местонахождении были обнаружены кости краниального и посткраниального скелета от разных особей мамонтов. Все найденные здесь бивни, по словам авторов находки, были небольшого размера.
|
Рис. 7. Местонахождение «Селлях», правый берег р. Селлях, верхнее течение, Усть-Янский улус РС(Я): I.-карта; II.-схема геологического разреза (см): 1 – дерн; 2 – суглинок (рыхлый); 3 – супесь; 4 – суглинок (плотный); 5 – остатки ископаемых млекопитающих; 6 – остатки растительности |
Из общей выборки остеологического материала преобладали остатки M. primigenius. Остатки других представителей мамонтовой фауны представлены единично и фрагментарно – E. lenensis, Bison sp., C. antiquitatis.
Высота берегового обрыва на местонахождении Селлях около 9-10 м, протяженность 40-50 м (рис. 8.I.). Обнажение образовалось около года назад, в первое время размеры обнажения были намного меньше (около 10 м). В последующем, вследствие термоабразионных процессов размеры местонахождения расширились (рис. 8. II.).
На местонахождении обнаружено 62 кости и фрагментов костей мамонтов. У многих трубчатых костей отсутствуют эпифизы. Практически все найденные кости имеют сильные повреждения абиотического характера. Результаты исследований остеологического материала и тафономии местонахождения показывают, что накопление было одно - или несколько-сезонным по сравнению, к примеру, с Берелехским «кладбищем» мамонтов, где накопление костного материала происходило в течение многих лет (Верещагин, 1977). Все костные остатки были найдены в радиусе 10-5 м и на глубине около 5-6 м, немногим выше уровня воды в реке, в одном горизонте (см. рис. 8. II.). Следов обработки костей древним человеком не обнаружено.
|
Рис. 8. Общий вид и схематическое расположение остеологического материала: I.- обнажение на правом берегу р. Селлях: II.- продольный разрез местонахождения и схема расположения костей. 1- урез речной воды; 2- предполагаемая граница расположения костных остатков; 3- расширенный вследствие термообразии участок местонахождения без костных остатков; 4- современный слой; 5- суглинок светло-серого цвета; 6- супесь, с прослойками светлого песка; 7- суглинок темно-серого цвета, костеносный горизонт; 8-9 - тонкий лед, вертикально пронизывающий почвенные горизонты; 10- уровень речной воды |

Размеры фрагментов черепов (рис. 9. I) с сохранившимися мыщелками уступают размерам таковых половозрелых самцов из других местонахождений (табл. 5) (Maschenko, 2002; Аверьянов, 1994; Тихонов, 1996; Портенко, 1951; Герц, 1902).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
