
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таблица 18.
Промеры первых шейных позвонков (atlant)
№ | Промеры (мм) | СГ8 | СГ9 | СГ10 | СГ11 | р. Моховая ЗИН 31736 40-50 лет | Гыданский п-ов, р. Юрибей; ПИН 3941 12-14 лет | Таймырский п-ов; ЗИН, 45-50 лет [1] |
1. | Ширина позвонка в области реберно-поперечных отростков | 340 | 320 | - | - | 386 | 275 | 368 |
2. | Расстояние между краями верхней и нижней дуг | 170 | 168 | 55 | - | 190 | 171 | 242 |
3. | Длина гребня верхней дуги | 56 | 67 | 40 | 67 | 76 | - | - |
[1] - Гарутт, 1954
Лопатка мамонта №СГ-12 (левая). Передний и задний края лопатки частично разрушены, как и клювовидный отросток. Ювенильная лопатка №СГ-13 (правая) коричневого цвета, клювовидный отросток обломан. Фрагмент лопатки №СГ-14 (левая) палевого цвета. Фрагмент дистальной части лопатки №СГ-15 (левая) палевого цвета. Фрагмент дистальной части лопатки №СГ-16 (левая) коричневого цвета. Ювенильная лопатка №СГ-17 (правая) коричневого цвета, передний и задний края разрушены. Фрагмент лопатки №СГ-18 палевого цвета, передний и задний края, а также клювовидный отросток разрушены. Фрагмент проксимального конца левой лопатки №СГ-19, цвет серый (табл. 19). Приблизительный индивидуальный возраст лопаток мамонтов варьирует от 12-14 до 40 лет.
Таблица 19
Промеры лопаточных костей (scapula)
Промеры (см). | Обр. с Мус Хая | Юрибейский мамонт, Гыданский полуостров ПИН 3941 12-14 лет. | р. Березовка, ЗИН 5316 40-50 лет |
Длина сочленовной впадины | 13,0-19,6 | 15,3 | 19,1 |
Ширина сочленовной впадины | 10,9-7,0 | 8,8 | 10,2 |
Плечевая кость №СГ-20 (правая) бордового цвета. Проксимальный эпифиз отсутствует в связи с тем, что был не приросший. Возрастные швы на дистальном конце полностью облитерированы. Правая плечевая кость №СГ-21 ярко-коричневого местами палевого цвета. Проксимальный эпифиз выпал связи с тем, что был не приросший. Возрастные швы дистального конца полностью облитерированы. Плечевая кость (правая) №СГ-22 палевого цвета. Проксимальный и дистальный эпифизы сохранились. На проксимальном конце швы полностью не облитерированы. Левая плечевая кость №СГ-23 палевого цвета. Проксимальный конец обломан в середине диафиза вследствие механического воздействия (абиотического характера). Ювенильная плечевая кость мамонта №СГ-24 коричневого цвета (левая). Проксимальный и дистальный эпифизы выпали связи с тем, что были не приросшие. Левая плечевая кость палевого цвета №СГ-25 (левая). Проксимальный и дистальный эпифизы выпали вследствие их не приростания к диафизу. Левая плечевая кость №СГ-26 палевого, местами коричневого цвета. Эпифизы выпали. Фрагмент левой плечевой кости №СГ-27. Сохранился бугор латерального над мыщелка. Цвет темно-коричневый. Левая плечевая кость №СГ-28 светло коричневого, местами темно-коричневого цвета. Эпифизы выпали вследствие их не приростания. Левая плечевая кость палевого цвета №СГ-29. Эпифизы сохранились. На проксимальном конце швы полностью не облитерированы. Результаты сравнения представлены в таблице 20, где примерный индивидуальный возраст исследуемых экземпляров около 20-30 лет.
Таблица 20.
Промеры плечевой кости (humerus)
Промеры, см | Humerus СГ-19 (прав.) | Humerus СГ-20 (прав.) | Humerus СГ-21 (прав.) | ПИН 4353-630 [1] | ЗИН 31736 [2] |
Общая длина | 73,0 | 72,0 | 73,5 | 56,0 | 97,4 |
Дистальный конец: передне-задний/ поперечный | 23,0/18,0 | 23,0/27,0 | 26,5/19,0 | -/18,0 | -/27,5 |
Проксимальный конец: передне-задний/ поперечный | 25,5/х | - | - | -/14,0 | -/- |
Медиальная част диафиза: передне-задний/ поперечный | 11,0/9,0 | 11,0/12,0 | 11,5/11,5 | -/- | /11,6 |
Возраст | 20-30 | 2030 | 20-30 | 6-7 лет | 40-50 лет |
[1] - Maschenko, 2002; [2] - Аверьянов, 1994
На местонахождении найдено 13 экземпляров тазовых костей, которые имели сильные повреждения абиотического характера. Из общей выборки по размерным показателям 5 экземпляров принадлежат молодым особям. Некоторые кости были покрыты пятнами вивианита, которые обычны в низинных торфяных болотах.
Расчет количества особей проводился по соотношению и количеству левых и правых костей конечностей, а также наличие таких костей краниального и посткраниального скелета как нижние челюсти и тазовые кости. Из общей выборки для определения возрастного состава находок были использованы 5 нижних челюстей с сохранившимися зубами, а также 2 изолированных зуба. Кроме того, в этих же целях были использованы 10 плечевых и 10 бедренных костей.
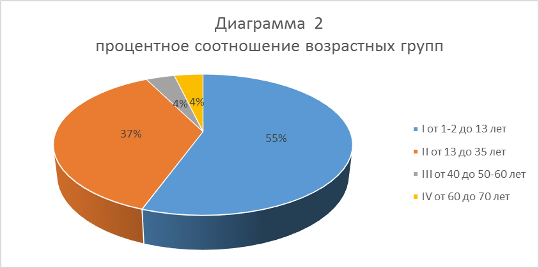
По возрастным показателям костей конечностей (характер срастания эпифизов к диафизу и общие размеры) и зубов было выявлено 4 возрастные группы: I - от 1-2 до 13 лет; II от 13 до 35 лет; III от 40 до 50-60 лет; IV 60-70 лет.
В группу I подпадают кости конечностей с не сросшимися дистальными и проксимальными эпифизами: плечевые - №СГ-24, №СГ-25, №СГ-26, №СГ-27, №СГ-28; бедренные - №СГ-43, №СГ-44, №СГ-45, №СГ-46, №СГ-47, №СГ-48, а так же нижние челюсти с генерацией зубов dp3-dp4, dp4-m1 - №СГ-3, №СГ-4.
Во вторую группу включены кости конечностей с не приросшими проксимальными концами, но с приросшими дистальными эпифизами: плечевые - №СГ-20, №СГ-21, №СГ-23; бедренные - №СГ-49, №СГ-50, №СГ-51, №СГ-52. В эту же группу включены две плечевые кости - №СГ-22, №СГ-29 относительно не больших размеров с обоими приросшими эпифизами, возрастные швы которых на проксимальных концах полностью не облитерированы. По-видимому, эти кости относятся к самкам, у которых прирастание эпифизов происходит раньше (Roth, 1984). Сюда же включены два изолированных зуба (№СГ-6, №СГ-7) и нижняя челюсть с генерацией зубов m2-m3 (№СГ-2).
В возрастную группу III включена нижняя челюсть с генерацией зубов m3 (№СГ-5).
В группу IV включена нижняя челюсть №СГ-1, с почти полностью стертой последней генерацией зубов М3.
Таким образом, количество мамонтов входящих в возрастную группу I составляет 5 особей – 55,5%, группа II включает 3 особи – 37% и группы III -3,7% и IV – 3,7% по одной. В результате исследуемый остеологический материал принадлежал 13 разновозрастным особям мамонтов от 2 до 70 лет (диаграмма 2).
Одним из признаков определяющих половую принадлежность является симфизный отросток на нижней челюсти. Симфизный отросток у самцов мамонтов крупнее и массивнее чем у самок (Гарутт, 1992). Таким образом, экз. №СГ2 с крупным симфизным отростком мог принадлежать самцу. Экземпляры №СГ1, №СГ4, №СГ5, у которых менее выражен симфизный отросток принадлежат самкам. Экз. №СГ3 еще не достиг возраста проявления полового диморфизма (10-18 лет) (Мащенко, 1992). Определителем видовой принадлежности у хоботных является последний коренной зуб – m3. Для M. primigenius частота пластин на 10 см должно быть не менее 8-9. Частоту пластин определяет характер питания, т. е. жесткая трава и ветки или мягкие сочные листья. Судя по частоте пластин M. primigenius питались травой, ветками, листьями кустарничков и кустарников, хвоей лиственницы, мхами. Но при этом первостепенное значение в рационе имели злаковые, осоковые и разнотравье (Томская, 2000). Таким образом, изученные нами нижние челюсти принадлежат M. primigenius.
Как описывалось выше в 2001 недалеко от МусХая ниже по течению р. Яна была открыта стоянка древнего человека. На этой стоянке были обнаружены изделия из костей и «кухонные» остатки млекопитающих мамонтовой фауны: C. lupus, A. lagopus, U. arctos, G. gulo, P. spelaea, M. primigenius, C. antiquitatis, Equus sp., R. tarandus, B. priscus и O. pallantis.
Большое скопление остатков животных мамонтовой фауны образуется в течение длительного времени. Примером для данной интерпретации является захоронение мамонтов на берегу р. Берелех. На местонахождении Берелехское «кладбище» мамонтов были обнаружены около 10 тысяч костей (и это не предел), принадлежащих 150 особям взрослых самок, детенышей и молодых мамонтов. (1977) и и др. (2010) объясняют это обстоятельство особенностями поведения и среды обитания животных. Их жизнь, поиск пищи, и сезонные миграции происходили в основном по долинам рек. На опасных участках речных долин слабые мамонты, попавшие в естественную природную ловушку погибали, и из года в год, вероятно, накапливались на этом месте, образуя «кладбище» мамонтовой фауны.
(2011) придерживается версии об антропогенном происхождении захоронения мамонтовой фауны на местонахождении Мус Хая, как результат охоты древнего человека. Данное утверждение основывается автором на соответствие радиоуглеродных датировок полученных со стоянки и в захоронении животных мамонтовой фауны, а также обнаружение костных остатков со следами деятельности человека. Хотя прямых доказательств охоты человека на мамонтов здесь не обнаружено.
Количество костного материала на массовых захоронениях мамонтов исчисляется десятками тысяч. Мус Хая не является исключением, видимо, накопление костей животных происходило по схеме берелехского «кладбища» мамонтов, т. е. не далеко (вверх по течению) могла находится миграционная тропа животных, которая пролегала через древнюю реку Яна. С другой стороны, наличие стоянки древнего человека не далеко от захоронения мамонтов также можно объяснить тем, что человек селился возле естественных захоронений для сбора необходимого ему материала для строительства и изготовления орудия труда (кости, бивни и т. п.).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
