
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На вентральной поверхности основания хобота имеется впадина шириной 50.0 мм и длиной 170 мм, локализованная на уровне окончания неба и начала вентральной поверхности хобота (рис 20). Латерально по отношению к этому углублению расположены складки кожи, окаймляющие альвеолы бивней. Диаметр отверстий альвеол бивней 30.0 х 35.0 мм. В основании хобота имеется слой подкожной соединительной ткани. Дистальнее, приблизительно от уровня проксимальной трети от длины хобота она сменяется тонкими и слабо дифференцированными друг от друга продольными мышечными волокнами. Толщина кожи на проксимальном конце хобота - 2,13 мм.
На расправленном хоботе ширина расширения достигает 140 мм. На полностью расправленном хоботе, в его нижней трети латеральные складки растягиваются в поперечном направлении, формируя расширение. Данные о локализации расширения на хоботе представлены в таблице 17.
Таблица 17.
Размеры "расширения" хобота у мамонта Юка.
Промеры (мм) | Самка мамонта Юка. Экз. ОЮ № 1 |
Диаметр хобота на уровне альвеол бивней/в средней части хобота (приблизительного верхняя граница "расширения")//над основанием вентрального пальцеобразного концевого отростка | 140.0-150.0/80.0//75.0 |
Максимальная длина/ширина "расширения" хобота | 300.0/140.0 |
Длина от верхней/нижней границы "расширения" хобота до дистального конца хобота | 480.0/180.0 |
Ширина хобота на уровне проксимальной/дистальной границы "расширения" | 80.0/75.0 |
Расширение сохраняет форму в расправленном состоянии, при искусственном сжатии хобота в продольном направлении оно незначительно меняет форму и его ширина становится меньше. В области расширения латеральные складки кожи наиболее толстые и загибаются вентрально. Это расширение напоминает капюшон кобры. Сходство дополняется тем, что при полностью расправленных латеральных продольных складках, форма поперечного сечения хобота изменяется с овальной, у проксимального конца хобота на эллипсовидную в районе расширения/капюшона у дистального конца, на уровне оснований концевых пальцеобразных отростков снова поперечное сечение снова становится овальным.
Продольные мышечные волокна хобота на уровне поперечного расширения чередуются с незначительно наклоненными относительно продольной оси хобота мышечными пучками, что говорит об особом типе подвижности мускулатуры в области поперечного расширения хобота. Видимо, от поверхностного слоя мышц берут начало поперечные мышечные пучки. На всем протяжении хобота в поверхностном слое мышц нет кольцевых мышечных пучков. Возможно, что указанные поперечные и кольцевые мышцы находятся в более глубоких слоях мускулатуры хобота. На данном этапе исследований их препарирование и изучение не проводилось.
Функциональное назначение расширения хобота, обнаруженное у самки молодого мамонта Юка, может быть интерпретировано по-разному. Нами рассмотрены две гипотезы, имеющие адаптационное объяснение.
Одной из гипотез может быть использование расширения концевой части хобота для согревания кончика хобота.
На отсутствие шерсти на концевых пальцеобразных выростах указывают такие косвенные данные, как рисунки людей верхнего палеолита на стенах пещер в Западной Европе. На двух рисунках поздней поры верхнего палеолита из Дордони конец хобота с пальцеобразными выростами четко изображается без шерсти, а вентральная поверхность хобота покрыта шерстью значительно более короткой на дистальной части хобота, по сравнению с вентральной поверхности его проксимального конца (рис. 25).
| Рис. 25. Изображения мамонта человеком верхнего палеолита на стенах пещер в Западной Европе. 1 - Пещера Фонт-де-Гом. Дордонь, Франция. Около 14 тыс. лет; 2 - Пещера Комбарелль, Дордонь, Франция. Около 15 тыс. лет. |

Зимой, мамонты тратили на сбор корма не меньше времени, чем современные африканские слоны в засушливый сезон (Shoshani et al., 1991; Haynes, 1991). Это предполагает, что пальцеобразные концевые отростки оставались открытыми и контактировали с охлажденными до температуры воздуха растениями и почвой более 15-16 часов в сутки. При таком охлаждении необходимый обогрев пальцеобразных отростков мог производится в расширение дистальной трети хобота. Как и современные слоны, мамонт закручивал хобот в «спираль» и концевая часть хобота дополнительно прикрывался боковыми складками расширения, как бы образуя меховую «варежку» (рис. 26).
|
Рис. 26. Схематическое изображение спирального закручивания хобот. А - у современного слона; б – у шерстистого мамонта. 1- расширение на концевой части хобота; 2 – при спиральном скручивании кончик хобота мамонта попадает между складками его расширения, оказываясь прикрытым меховой частью |
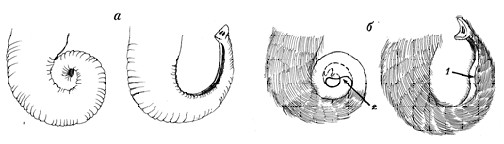
Другая гипотеза объясняющая формирование концевого расширения хобота может быть связана с поеданием шерстистыми мамонтами снега при нехватке, не замерзшей воды. Одним из первых идею о нехватки, не замерзшей воды для шерстистого мамонта в условиях холодной и сухой зимы позднего плейстоцена высказал (1979). Прямых доказательств поедания снега мамонтами зимой нет, но не рассматривать эту возможность для M. primigenius нельзя.
Современные растительноядные млекопитающие нуждаются в постоянном водопое и пополнении воды в организме и количество необходимой воды не зависит от времени года. Высокое содержание воды в желудке растительноядных животных необходимо для переваривания растительной пищи и нормального существования одноклеточных симбионтов, обеспечивающих сбраживание и расщепление клетчатки. Кроме того, всасывание расщепленных питательных веществ требует формирования мембран и пленок основным компонентом которых является вода. Изменение количества воды в отделах желудка и прилегающих отделах тонкого кишечника принципиально для выживания этих животных.
У косуль (Capreolus capreolus Linnaeus, 1758) в любой сезон года доля воды в содержимом желудка составляет не менее 80%. При недостатке воды зимой косули вынуждены поедать снег, и тем больше, чем меньше воды содержится в пище. Однако снег почти лишен солей, и на его расплавление требуются дополнительные, весьма существенные энергозатраты. Поэтому, в нормальных условиях и при изобилии естественной влажной пищи косули и другие дикие копытные снег практически не употребляют, как и сухие корма, влажность которых составляет всего 12-22% (Данилкин, 1999). В России, с ее суровым климатом и многоснежьем, зимой диким копытным не хватает, не только пищи, но и воды. При кормлении европейских косуль сухими кормами вволю (при наличии свободной воды они, тем не менее, ежемесячно теряют до 9,4% массы тела (Приедитис, 1980).
Лось (Alces alces Linnaeus 1773) зимой явно избирает для питания наиболее влагоемкий корм и, очевидно, сокращает выделение влаги с мочой (Данилкин, 1999). Осенью и зимой для многих копытных основным источником пищи становятся зимнезеленые полукустарнички, древесные и наземные лишайники, осоки, хвощи (Романов, Беляков, 1980; Смирнов, 1987) и побеги хвойных пород, т. е. растения, содержащие наибольшее количество воды (Данилкин, 1999). На северо-востоке Сибири при низких температурах лось копает из-под снега осоки и хвощи, съедая до 6,5 кг влажного подснежного корма (Домнич, 1982). Побеги хвойных деревьев и зимнезеленых полукустарничков содержат в тканях наибольшее количество незамерзшей воды. Потребляя эту воду, животные экономят энергию при ее нагревании до температуры своего тела. Очевидно, животным в энергетическом отношении невыгодно использовать в пищу растения, не содержащие незамерзшую воду, или использовать в качестве источника воды снег (Данилкин, 1999).
Можно предположить, что количество воды необходимое шерстистому мамонту зимой было меньше, чем летом, но для половозрелой особи с весом в 3.5-4 тонны это количество, видимо, не может быть меньше, чем 100-120 литров в сутки. Современные слоны, расходуют часть воды организма для регулирования теплоотдачи и охлаждений организма и суточная норма воды для L. africana составляет 150 л (200 л для крупных особей) (Haynes, 1991). Можно предположить, что при отсутствии потовых желез на коже мамонт мог зимой "экономить" воду подобно лосям, сокращая ее выделение с мочой. Вместе с тем, вероятнее всего, количество доступной зеленой массы в условиях перегляциальной, сухой "тундростепи" было очень мало и значительную часть пищи мамонта составляли растения закончившие вегетацию (прошлогодние травянистые растения и "спящие" побеги кустарников и деревьев).
Все перечисленные для растительноядных млекопитающих особенности использования воды и вероятный уровень потребления воды мамонтом говорят о том, что замерзающих водопоях мамонтам приходилось поедать снег для восполнения потери воды. Одним из компенсаторных механизмов потери воды у шерстистого мамонта могло быть расщепление накопленного жира в организме. Основным продуктом такой физиологической реакции является образованию воды. Для шерстистого мамонта достоверно установлено накопление запасов жира. Оно отмечено как для детенышей этого вида (Мащенко и др., 2012; Мащенко и др., 2013), так и для взрослых особей (Верещагин, 1979; Верещагин, Тихонов, 1990). Однако, исследования процесса расщепления запасов жира у двугорбого верблюда (Camelus bactrianus Linnaeus 1773) показывает, что поступление воды освободившейся при расщеплении жира не покрывает всей потери воды при ее нехватке (Барабаш-Никифоров, Формозов, 1963). Поэтому для шерстистого мамонта компенсация нехватки воды только за счет расходования запасов жира маловероятна и более вероятно восполнение ее дефицита за счет поедания снега.
Несмотря на вероятность использования мамонтом снега зимой при отсутствие не замерзшей воды, существует ряд достаточно жестких физиологических ограничений на поедания снега этими млекопитающими. Наиболее существенным ограничением потребления снега шерстистым мамонтом является угроза нарушения теплового баланса организма, такой дисбаланс мог возникнуть из-за слишком больших энергетических затрат необходимых только на растапливание минимум 2-3 м3 снега в течение суток для получения 100-120 литров воды необходимой мамонту.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
