
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ГЛАВА 3. ШЕРСТИСТЫЙ МАМОНТ - ГЛАВНЫЙ ИНДИКАТОР МАМОНТОВОЙ ФАУНЫ В ПОЗДНЕМ НЕОПЛЕЙСТОЦЕНЕ
3.1. Шерстистый мамонт
Мамонт это древний слон, обитавший в позднем неоплейстоцене и вымерший в начале голоцена. Мамонт был всецело адаптирован к холодному климату. Он был покрыт густой и длинной шерстью, которaя свисая с брюха и ног, образовывала подобие шерстяной «юбки». Тело мамонта было овальной формы, уши были небольших размеров, как и хвост. Замечательным приспособлением для добывания пищи стали два видоизмененных резца - бивни мамонтов. Они использовали их для добывания пищи из-под снега и турнирных битв за самку в период спаривания (Верещагин, 1979, Плотников, 2010).
В течение всего периода существования мамонты претерпевали некоторые морфологические изменения, т. е. адаптивные изменения, приспосабливавшие их к окружающей среде. К подобным изменениям относится размеры мамонтов. В конце XX века на острове Врангеля были обнаружены кости мамонтов некрупных размеров, результаты радиоуглеродных датировок показали, что они существовали около 3700-4000 лет назад (Vartanyan et al., 1993; Вартанян, 2007). Кроме того, автор во время экспедиции на о-ве Врангеля, исследовал бивень пожилой самки мамонта, реконструированная высота тела которой оказались довольно небольшой (Плотников, 2012).
По мнению некоторых ученых (Верещагин, 1979; Тихонов, 2005; Вартанян, 2007) к закату своего существования мамонты измельчали в размерах. Однако, по мнению (1976) мамонты претерпевали изменения размеров тела несколько раз, т. е. столько раз, сколько было оледенений и межледниковий. В холодные эпохи (зырянское, сартанское время) мамонты увеличивались в размерах, а в теплые периоды (казанцевское, каргинское) животные наоборот уменьшались в размерах.
Этому факту свидетельствуют находки некрупных мамонтов на острове Врангеля, датируемых теплым голоценовым периодом (Вартанян, 2007), а также некоторые находки крупных мамонтов на местонахождениях «Волчья грива» (Новосибирская область) 14200 ±150 л. н. (СО АН-78) – сартанское оледенение (Шпанский, 2007).
Автором предлагаемой работы измерялись кости некоторых скелетов мамонтов, для проведения самостоятельных исследований и сравнения с данными полученными другими авторами. Как показали результаты измерений, очень крупные особи мамонтов встречались на севере Восточной Сибири как во время каргинского потепления (Ленский (выс. в холке 308 см) и Тирехтяхский (выс. в холке ок. 290 см) мамонты), так и во время сартанского оледенения (Аллаиховский мамонт (выс. в холке ок. 290 см). Сравнительно мелкие особи мамонтов обитали и во время каргинского термохрона (Березовский, выс. в холке 265 см) и в период сартанского криохрона (Таймырский, выс. в холке 250 см); берелехские мамонты (Барышников и др., 1977). Таким образом, можно констатировать, что мамонты севера Восточной Сибири, в том числе и Яно-Индигирской низменности, во второй половине позднего плейстоцена отличались большой вариабельностью в размерах, от исключительно крупных особей, близких к максимальным для вида, до сравнительно мелких, причем, как во время потепления, так и во время похолодания.
По сравнению с современными слонами, у мамонта были более короткие конечности. Это объясняется характером добычи корма, мамонт питался подножным кормом, в то время как слоны предпочитают питаться ветками и листьями деревьев (Верещагин, 1979).
Мамонтам, как растительноядным млекопитающим крупных размеров, ежедневно требовалось, по-видимому, от 250 до 350 кг корма (Тихонов, 2005). Добыть его можно было только в долинах рек, у озер и болот. Вероятно, мамонты летом заходили далеко на север в богатую травой тундростепь, а зимой откочевывали к югу, как это делают современные северные олени.
Ареал обитания мамонтов наряду с изменением окружающей среды менялся, т. е. во времена пиков численности, мамонты обитали по всей Евразии и отчасти в Северной Америке. Затем, по мере изменения климатических и ландшафтных условий, их ареал тоже менялся, становился прерывистым и локализованным на относительно небольших участках.
На пике численности ареал обитания мамонта покрывал практически всю Евразию от Берингии до Британских островов и часть северной Америки. Территория Яно-Индигирской низменности несомненно полностью перекрывается ареалом распространения мамонтов, шерстистых носорогов и других животных мамонтовой фауны. По данным (1995) и и др. (2001), около 50 – 20 тыс. л. н. южная граница обитания мамонтов проходила по 50° северной широты, которая на стыке позднего неоплейстоцена и голоцена (12 тыс. л. н.) сместилась на 70° с. ш (рис. 3).
|
|
| Рис.3. Ареал шерстистого мамонта в разных промежутках времени. Голубым цветом обозначена территория Яно-Индигирской низменности. |
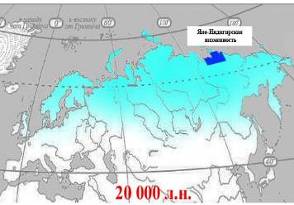

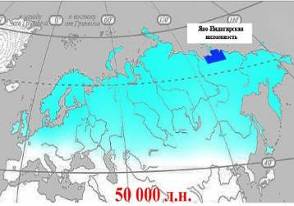
Таким образом, территория Яно-Индигирской низменности беспрерывно являлась местом обитания животных мамонтовой фауны, вплоть до начала голоцена.
Исследуемая территория является одним из тех мест, где часто находят не только костные остатки, но и хорошо сохранившиеся туши вымерших животных (рис.4).
|
Рис. 4. Местонахождения M. primigenius на территории Яно-Индигирской низменности: 1- Берелехское «кладбище» мамонтов; 2 - Максунуохский мамонт; 3 – Юкагирский мамонт; 4 – Хромский детеныш мамонта; 5 – Мамонт Юка; 6 – Хапташинский яр; 7 – р. Аччыгый Аллаиха; 8 – р. Омолой; 9 – р. Мылахчын; 10 - Мус Хая; 11 – Тимирдях хайа; 12 – Куччугуй куегулур; 13 – Бургуат; 14 – Киэнг-Юряхский мамонт; 15 - Селляхские мамонты; 16 – оз. Булгунньяхтах, 17 – местонахождение «Ыгыанньа»; 18 – р. Буор Юрях. |

Мамонты с Берелехского «кладбища»
В 2008 г. нами в составе экспедиционного отряда ИПЭС под руководством проведены исследования современного состояния особо охраняемой территории «Берелехское кладбище мамонтов».
На время работ отряда стоял высокий уровень воды в реке, примерно на 2 м выше, чем в 1970 году. Весь пляж обнажения находилась под водой, поэтому на берегу обнаружили очень мало костей. Места раскопок 1970 и 1972 годов полностью разрушены солюфлюкционно-оползневыми процессами. На их месте в момент обследования образовалась широкая ниша, покрытая дерном и заросшая травяным покровом. На ранее обследованной части местонахождения костных остатков не обнаружено. Выше по течению реки от места раскопок образовалось новое широкое обнажение мощных жильных льдов белого и сероватого цветов. Льды, уплотненные с вертикальной полосатостью и пузырьками. Края слоев покровных лессовидных суглинков у контакта с жильным льдом изогнуты вверх, что является результатом сжатия жильного льда. Ширина жильных льдов местами достигает 6-7 м.
В 1970 г. выше основного места скопления костей в 15 м от берегового обрыва реки была вырыта яма, куда сложили несколько сот костей плохой сохранности и закопали. В этом году обнаружили, что яма с костями при разрушении берега обвалилась в реку. Сохранилось немного костей мамонта, в основном, обломки позвонков ребер, зубов. В результате долгого нахождения в аэробной среде эти кости подверглись сильному выветриванию.
Берелехское кладбище могло, накопится за счет гибели подростков и молодых особей, так и за счет небольших групп. Отсутствие особо крупных особей в Берелехе может объясняться их большой осторожностью и силой. Вероятно, не далеко верх по течению р. Берелех пролегала миграционная тропа мамонтов, которая пересекала реку. Если принять концепцию гибели мамонтов в полыньях, на непрочном льде и в половодьях, а также учитывать значительную мощность костеносного горизонта, то можно считать, что гибель зверей здесь и накопление их остатков происходили длительно и последовательно на протяжении многих сотен лет (Отчет НИР…, 2008).
Детеныш мамонта с р. Хрома
Замороженный труп детеныша мамонта (M. primigenius), был найден в октябре 2008 г. жителем пос. ебедевым. Раскопки были организованы ФГНУ ИПЭС под руководством и личным участием автора. Местонахождение расположено на склоне правого берега р. Хрома в ее нижнем течении (Аллаиховский улус). Труп детеныша мамонта обнаружен на высоте около 9 м от уровня воды и приблизительно в 2 м ниже кровли берегового склона. Данные геологической съемки разреза и строение аналогичных разрезов этого района Колымско-Индигирской низменности показывают, что верхняя часть разрезов до уровня 13,5 м состоит из льдистых отложений едомной свиты (Лазарев, Томская, 1987; Федоров и др., 1989). Отложения, непосредственно вмещающие труп детеныша, сложены лессовидными суглинками, супесями и жильными льдами. Труп in situ находился в вертикальном положении, с вытянутыми вперед передними и отогнутыми назад задними ногами (рис. 5а). Труп детеныша извлечен вместе с монолитом вмещающей его мерзлой породы весом около 350 кг и в мае 2009 г. доставлен в г. Якутск. Определение геологического возраста хромского детеныша по фрагменту ребра методом AMS, проведенное в Лаборатории университета г. Гронинген (Нидерланды) показало возраст более 50 тыс. лет.
Морфологические исследования этого детеныша проводились при нашем участии (Lazarev et al., 2010; Мащенко и др., 2012). До начала раскопок на голове и верхней части спины, выступавшие из мерзлых пород (от носового отверстия до уровня 11 грудного позвонка) песцами были съедены шкура и мягкие ткани. Пропорции хромского детеныша изменены продольным растягиванием туловища, которое вызвало разрыв трупа в средней части грудного отдела.
Передние и задние конечности сильно согнуты в локтевых и коленных суставах, деформированы и сведены друг с другом. Из-за деформации конечности выглядят непропорционально толстыми и короткими. В передней половине туловища отсутствуют внутренние органы грудной клетки: легкие и сердце. Диафрагма разорвана. Через разрывы кожи на правой и левой поверхностях тела видны внутренние органы передней части брюшной полости. Деформация и растяжение туловища, вероятнее всего, произошли сразу после гибели, либо само растягивание было травмой, вызвавшей гибель. Этот вывод можно сделать по явными следам растяжения шкуры на латеральных поверхностях туловища (рис. 5а, 5б). Сильное продольное растягивание туловища выражается в том, что его вертикальный и поперечный диаметры в области разрыва составляют 320 и 270 мм, соответственно (см. рис. 5а, 5б). Вместе с неровными краями разорванной шкуры растяжение мягких тканей в виде продольных лоскутов могло образоваться только при растягивании тела живого или только что умершего детеныша.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
