


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
1.10. Ca2+-флуоресцентные красители, возбуждаемые видимым светом
Применение Ca2+-индикаторов с целью измерения уровня концентрации ионов кальция предусматривает возбуждение флуоресцентного свечения видимым или УФ-светом. Индикаторы, возбудимые видимым светом, имеют ряд преимуществ относительно УФ-возбудимых (Indicators for Ca2+, www. ):
- Эффективное возбуждение свечения малой энергией при использовании большинства лазерных приборов, включая конфокальный сканирующий микроскоп. Более сильное поглощение красителей, что позволяет использовать меньшую концентрацию красителя, снижая тем самым влияние фототоксичности на живые клетки. Большее изменение интенсивности Ca2+-зависимой флуоресценции при регистрации транзиента Ca2+. Излучение в той области спектра, где более низкий шум клеточной автофлуоресценции от исследуемого препарата и малое рассеянное фоновое свечение. Совместимость с фотоактивируемыми зондами и другими УФ-поглощающими реагентами, что удобно при проведении многопараметрических измерений. Длинноволновые Са2+-индикаторы способны накапливать увеличение выбранного параметра для одновременного измерения прочих физиологических параметров, таких как рН и мембранный потенциал.
Кальциевые индикаторы разделяют на высокоаффинные и низкоаффинные.
Длинноволновый кальциевый индикатор Oregon Green 488 BAPTA 1-калиевая соль (Рисунок 4) относится к высокоаффинным красителям и является исключительно ярким красителем. Он легко возбудим светом видимого диапазона. Этот индикатор разработан в форме водорастворимой соли калия. Связываясь с кальцием, индикатор демонстрирует увеличение интенсивности флуоресцентного излучения при малом смещении длины волны. В настоящее время индикаторы Oregon Green используются при изучении динамики кальция в изолированных и неизолированных препаратах.
Поглощательная способность Oregon Green BAPTA-1 при 488нм составляет приблизительно 93% от своего пикового значения, в то время как поглощательная способность индикаторов fluo-3 и Calcium Green при 488нм составляет только 45% от их максимумов. Следовательно, Oregon Green 488 BAPTA более эффективно возбуждается на длине волны 488 нм, чем красители fluo-3 и Calcium Green (Indicators for Ca2+, www. ).

Рисунок 4 - Красители Calcium Green и Oregon Green 488 BAPTA с различным сродством к Ca2+
Спектральные свойства Oregon Green 488 BAPTA (Рисунок 5) позволяют использовать низкие концентрации красителя. Кроме того, квантовый выход комплексов Oregon Green 488 BAPTA и Calcium Green составляет приблизительно 0.7, по сравнению с 0.14 для fluo-3. Как и Calcium Green-1, Oregon Green 488 BAPTA-1 умеренно люминесцирует в растворе без Ca2+, а при насыщении Ca2+ его флуоресценция повышается приблизительно в 14 раз.

Рисунок 5 -. Ca2+-зависимый спектр излучения флуоресценции красителя Oregon Green 488 BAPTA-1
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
2 МАТЕРИАЛЫ И МЕТОДЫ
2.1 Объект исследования
Эксперименты проводили на лабораторных мышах обоих полов. Животных умерщвляли перерезкой сонных артерий под эфирным наркозом. Эксперименты проводили на изолированном нервно-мышечном препарате разгибателя уха levator auris longus (LAL) белой лабораторной мыши. Животных анестезировали и декапетировали в соответствии с требованиями этических норм по работе с лабораторными животными (директива совета ЕС 86/609/EEC от 01.01.01 года). Нервно-мышечный препарат LAL в силу своих морфологических особенностей хорошо применим для микроскопических исследований.
Небольшая толщина и малые размеры позволяют использовать оптику Номарского для визуализации нервных окончаний в режиме «светлое поле». Нервные ответвления хорошо визуализируются в обычном стереомикроскопе. Небольшие размеры LAL и малая толщина обеспечивают достаточную выживаемость препарата при длительной экспозиции с красителем. В LAL представлены в основном быстрые мышечные волокна. Изолированные препараты помещали в экспериментальную ванночку, через которую протекал раствор Рингера-Кребса следующего состава (ммоль/л): NaCl – 120.0, KCl – 5.0, CaCl2 – 2.0, MgCl2 – 1.0, NaHCO3 – 11.0, NaH2PO4 – 1.0, глюкоза – 11.0, рН раствора поддерживали на уровне 7.2-7.4. Для предотвращения мышечных сокращений во время проведения экспериментов добавляли в среду, омывающую препарат, мю-контоксин G III B, блокирующий натриевые каналы мышечного типа и предотвращающий распространение потенциала действия по мышце. Это позволяло проводить эксперименты в условиях приближенных к естественным без изменения концентрации кальция в физиологическом растворе.
2.2. Электрофизиологические исследования
Электрофизиологические эксперименты выполняли на диафрагмальном нервно-мышечном препарате мыши в условиях сниженного квантового состава. Пониженное содержание ионов кальция в среде и присутствие магния предотвращали мышечные сокращения в ответ на стимуляцию нерва(Са2+ 0.4 мМ, Mg2+ 6 мМ).. Стимуляцию двигательного нерва осуществляли прямоугольными импульсами длительностью 0.1 мс супрамаксимальной величины с интервалом 2 с. Регистрацию токов нервного окончания и ТКП производили с помощью одного экстраклеточного микроэлектрода, подведенного при визуальном контроле участку нервной терминали. Микроэлектрод с диаметром кончика 2.5-3.5 мкм, заполненный раствором Рингера или NaCl (0.5ммоль/л), имел сопротивление 2.0-4.0 МОм. Микроэлектрод изготавливали из трубок диаметром 1.5-1.8 мм, содержащих микрокапилляры, на полуавтоматической кузнице МЭ-2. Кончик микроэлектрода обламывали и оплавляли на специальной установке, изготовленной в нашей лаборатории.
Под визуальным контролем микроэлектрод подводили к терминали.
При определении квантового состава мы использовали методику выпадений [Castillo, et al 1954] , описанную выше, согласно которой квантовый состав определялся по формуле :
![]()
![]() , где m - квантовый состав
, где m - квантовый состав
2.3. Оптические исследования
Оценка концентрации внутриклеточного кальция и его изменения в ходе различных воздействий на кальциевый метаболизм осуществлялась с применением современных методов флуоресцентного анализа.
Визуальное наблюдение объекта и регистрацию быстрого изменения кальциевого сигнала осуществляли при помощи экспериментальной установки, построенной на основе микроскопа Olympus BX 51 оснащенной высокоскоростной чувствительной камерой RedShirtimajing NeruoCCD-smq camera (RedShirtimajing, USA). Камера позволяет фиксировать слабые флуоресцентные сигналы с частотой 2000 кадров в 1 с. (1 кадр в 0.5 мс). При разрешении 80*80 пикселей. Этого достаточно для того чтобы проводить регистрацию кальциевых ответов с хорошим временным и пространственным разрешением на синапсах теплокровных и холоднокровных животных.
В качестве источника освещения использовали монохроматор Polyhrom V фирмы Till Photonics, который был настроен на длину волны возбуждения красителя – 488 нм.
При загрузке использовали кальциевый краситель Oregon Green 488 BAPTA-1 Hexapotassium Salt, cell impermeant, в концентрации 10 мМ, разведенный на дистиллированной воде с добавлением ГЭПЭСА в концентрации 5мМ для стабилизации pH раствора на уровне 7.2 – 7.4
Для регистрации кальциевого транзиента применялась методика загрузки кальциевого красителя через культю нерва, подробно описанная в обсуждении результатов. Количественное представление изменения интенсивности флуоресцентного сигнала выражали через отношение относительного изменения флуоресценции к фоновому свечению ∆F/F0 [Shahrezaei et al., 2006]. F0 регистрировали в каждом эксперименте. Фоновое свечение даёт вклад в базовый уровень флуоресценции, но не влияет на относительное изменение (Рисунок 6). В соответствии с этими соображениями осуществляли обработку сигналов: вычитали из регистрируемого сигнала фоновую составляющую и представляли величину кальциевого транзиента как отношение ДF/F0 .
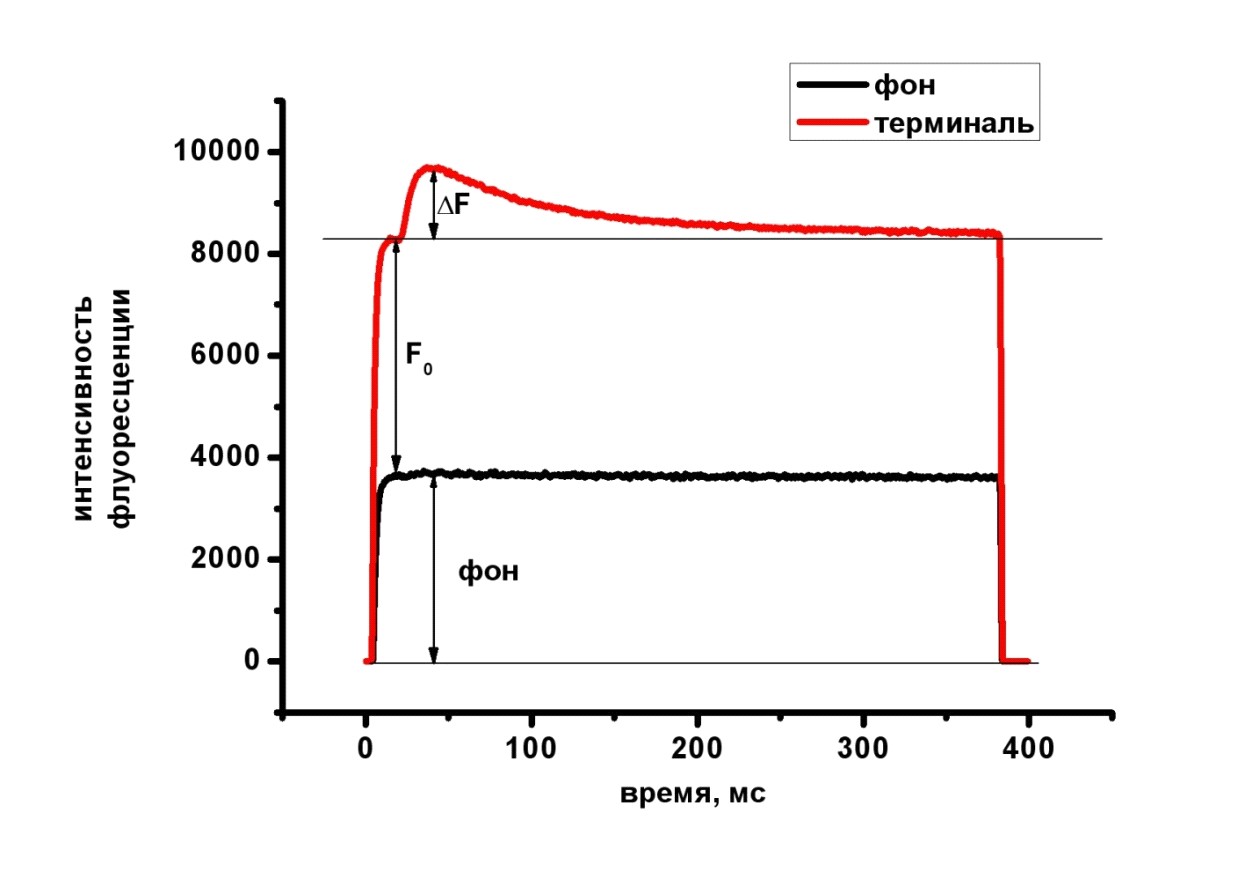
Рисунок 6. Количественное представление изменения интенсивности флуоресцентного сигнала
2.4. Статистическая обработка результатов
Для статистической обработки экспериментально полученных сигналов Ca2+-транзиента и электрофизиологических ответов использовали стандартные методы определения средних величин, стандартных ошибок, параметрический t-критерий Стьюдента для попарно связанных вариант. Достоверность различия средних значений определяли по стандартным критериям при P<0.05.
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
3.1. Влияние температуры на квантовый состав вызванных потенциалов концевой пластинки
Для анализа влияния температуры на величину квантового состава вызванных потенциалов концевой пластинки нами были проведены электрофизиологические эксперименты на изолированном нервно-мышечном препарате мыши. Мы регистрировали экстраклеточным методом вызванные потенциалы нервного окончания и вызванные потенциалы концевой пластинки. На рисунке 8 представлены Вызванные экстраклеточные потенциалы концевой пластики мыши, зарегистрированные при температуре 20![]()
![]() С.
С.
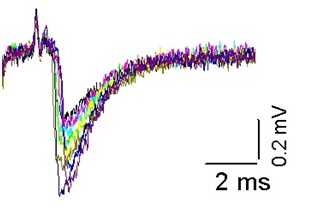
Рисунок 7. Вызванные экстраклеточные потенциалы концевой пластики мыши, зарегистрированные при температуре 20![]()
![]() С
С
Эксперименты проводились в условиях пониженного квантового состава при регистрации одноквантовых потенциалов пластинки. При этом оценивалось количество стимулов и количество одноквантовых сигналов. Квантовый состав рассчитывали методом выпадений, который описан выше в разделе методика.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
