


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
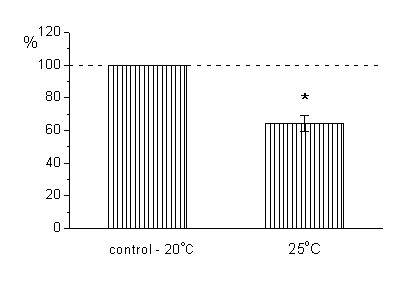
Полученные данные наглядно показали, что при повышении температуры от 20![]()
![]() С до 25
С до 25![]()
![]() С квантовый состав токов концевой пластинки уменьшился на 35.6± 4.8% (n=5)
С квантовый состав токов концевой пластинки уменьшился на 35.6± 4.8% (n=5)
Рисунок 8. Изменение квантового состава токов концевой пластинки мышцы мыши при увеличении температуры от контроля 20![]()
![]() С до 25
С до 25![]()
![]() С. Значения квантового состава на графике выражены в процентах от контроля. Различия достоверны, Р<0.05. Метод подсчета достоверности описан в методике выше. Абсолютные значения квантового состава изменились с 0,3458±0,03627 до 0,2284±0,03711
С. Значения квантового состава на графике выражены в процентах от контроля. Различия достоверны, Р<0.05. Метод подсчета достоверности описан в методике выше. Абсолютные значения квантового состава изменились с 0,3458±0,03627 до 0,2284±0,03711
Одним из возможных механизмов влияния температуры на величину квантового состава может быть влияние на пресинатпический уровень кальция во время стимуляции двигательного нерва. Для проверки этого предположения было решено оценить пресинаптический уровень кальция при различных температурах, используя методику анализа кальциевого транзиента.
3.2. Разработка методики доставки флуоресцентного красителя в двигательные нервные окончания теплокровных
Для проведения экспериментов по изучению кальциевого транзиента в нервных окончаниях мыши при различной температуре, необходимо было реализовать доставку кальциевого красителя в нервные окончания. Загрузку красителя выполняли на изолированном нервно-мышечном препарате разгибателя уха levator auris longus (LAL) белой лабораторной мыши. Нервно-мышечный препарат LAL в силу своих морфологических особенностей хорошо применим для микроскопических исследований.
Для загрузки кальциевого красителя использовали стеклянную микропипетку. Диаметр внутреннего отверстия получившейся микропипетки должен соответствовать диаметру выбранной для загрузки нервной ветки.
Под визуальным контролем с помощью стереомикроскопа стеклянную микропипетку подводили к выбранной нервной ветке и воском фиксировали на предметном стекле (Рис. 9). Непосредственно перед загрузкой красителя микроножницами рассекали нервную ветку. При помощи шприца нерв с небольшим объемом раствора засасывался в микропипетку. Для аппликации красителя на культю нерва использовали пластиковый микронаконечник собственного изготовления. Микронаконечник с раствором красителя вставляли в стеклянную микропипетку под визуальным контролем и при помощи шприца производили введение красителя (Рис 10.1).
Чашку Петри с закрепленной микропипеткой помещали в темное место на 2-3 часа.
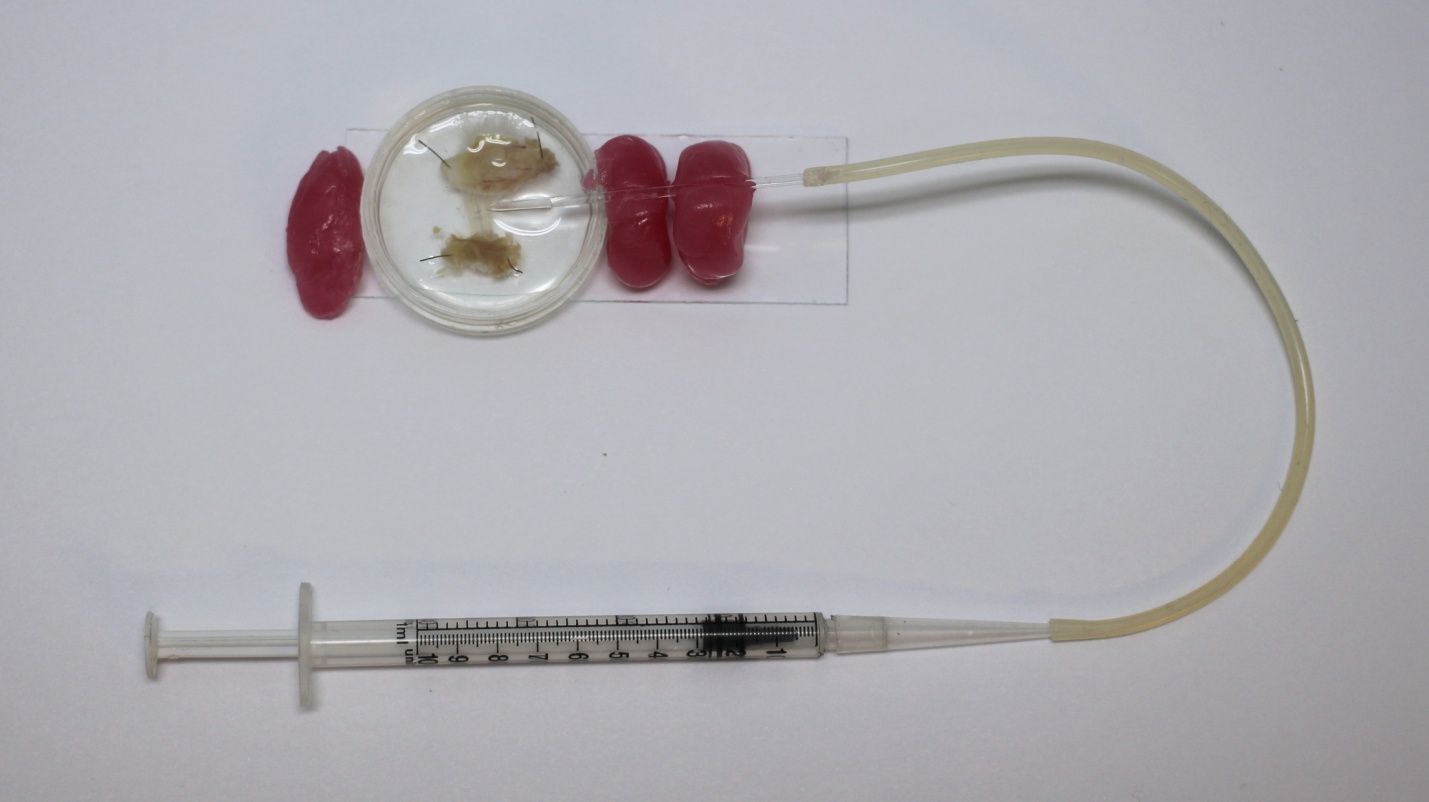
Рис. 9 Фотография препарата с закрепленной воском микропипеткой и шприцом для засасывания культя нерва.
В процессе инкубации препарата с красителем осуществляли проверку процесса загрузки красителя при помощи флуоресцентного микроскопа (Рис. 10.2).
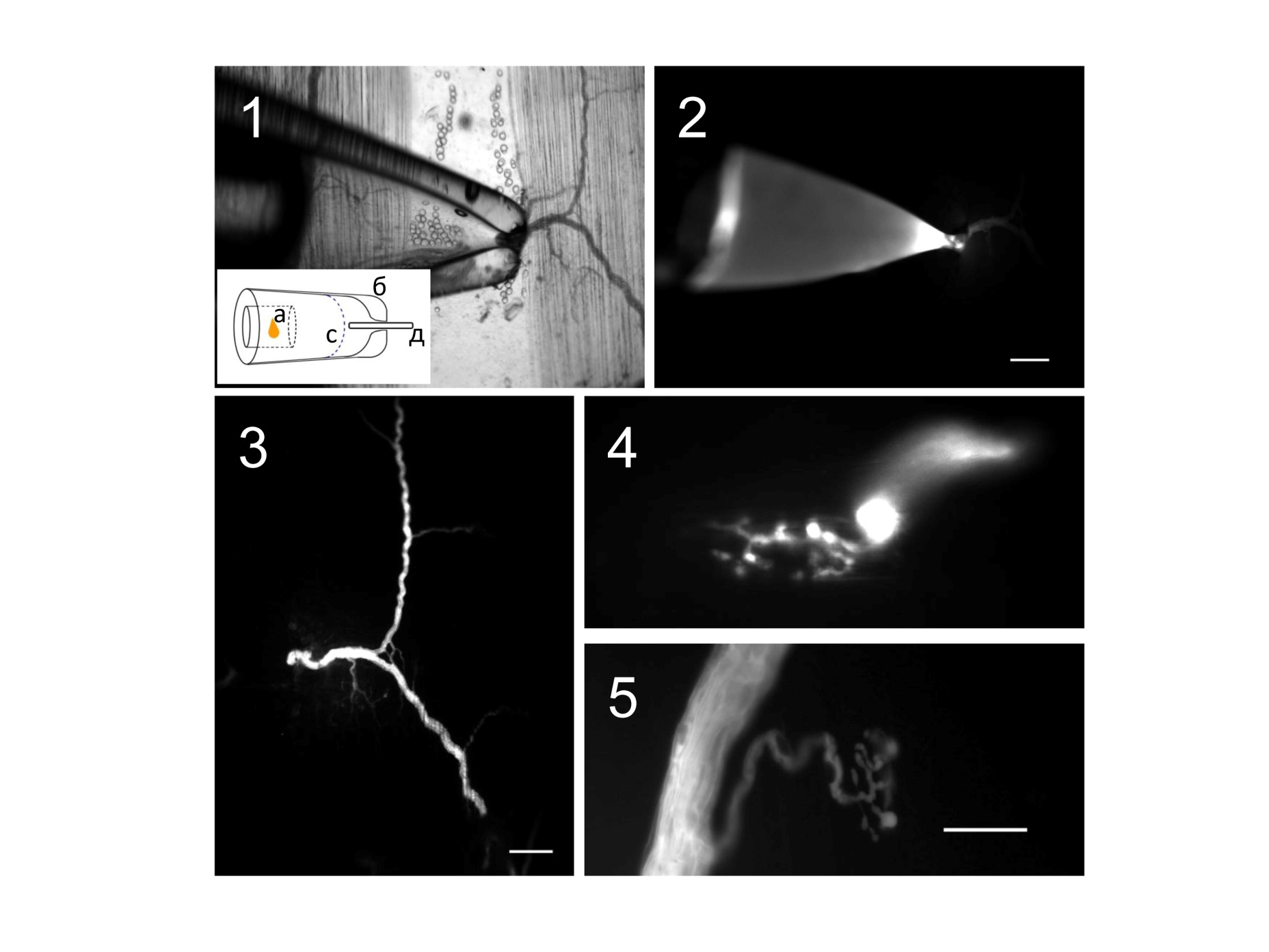
Рис 10. Микрофотографии загрузки нервных окончаний красителем Oregon Green 488 BAPTA-1. 1 – микропипетка с культей нерва и красителем в режиме освещения «светлое поле», на схеме показано изображение микропипетки (б) с введенным в нее микронаконечником (а) культей нерва (д) в растворе (с); 2 – та же область препарата, что и на 1 вкладке в режиме «флуоресценция», размерность шкалы 200мкм; 3 – сегмент нерва, загруженный красителем после трехчасовой инкубации, режим освещеения «флуоресценция», размерность шкалы 200мкм,; 4, 5 – нервные окончания загруженные красителем в режиме освещения «флуоресценция», размерность шкалы 20мкм
После процедуры загрузки красителя все проксимальные участки нерва и нервные ответвления с терминалями окрашивались флуоресцентным красителем (Рис. 10.3-5). Препарат перемещали в рабочую экспериментальную камеру и проводили процедуру регистрации кальциевого транзиента, описанного выше в разделе методика.
3.3 Влияние температуры на величину кальциевого транзиента в терминальных ответвлениях мыши
После загрузки кальциевого красителя препарат помещали в камеру и на экспериментальной установке, описанной в разделе методика, регистрировали флуоресцентные кальциевые сигналы(кальциевый транзиент). Регистрацию кальциевых сигналов и оценку величины амплитуды кальциевого транзиента проводили при изменении температуры раствора в диапазоне 20![]()
![]() С - 30
С - 30![]()
![]() С с шагом в 5
С с шагом в 5![]()
![]() С. На рис 11. Представлены зарегистрированные кальциевые транзиенты при варьировании температуры от 20 до 30 С0
С. На рис 11. Представлены зарегистрированные кальциевые транзиенты при варьировании температуры от 20 до 30 С0
Рис. 11 Изменение кальциевого транзиента пресинаптической терминали двигательного нерва LAL мыши при увеличении температуры от контроля 20![]()
![]() С до 30
С до 30![]()
![]() С с шагом в 5
С с шагом в 5![]()
![]() С. Количественное представление изменения интенсивности флуоресцентного сигнала выражали через отношение относительного изменения флуоресценции к фоновому свечению ∆F/F0 [Shahrezaei et al., 2006]. F0 регистрировали в каждом эксперименте.
С. Количественное представление изменения интенсивности флуоресцентного сигнала выражали через отношение относительного изменения флуоресценции к фоновому свечению ∆F/F0 [Shahrezaei et al., 2006]. F0 регистрировали в каждом эксперименте.
Было установлено, что при увеличении температуры до 25![]()
![]() С амплитуда кальциевого транзиента достоверно снижалась на 13.7±2.2% а при дальнейшем увеличении температуры до 30
С амплитуда кальциевого транзиента достоверно снижалась на 13.7±2.2% а при дальнейшем увеличении температуры до 30![]()
![]() С падала на 14.2±4.8. (рис 12) Таким образом наблюдаемое уменьшение квантового состава при увеличении температуры может быть связано с уменьшением пресинаптического уровня кальция, о чем говорит уменьшении кальциевого транзиента при увеличении температуры окружающей среды.
С падала на 14.2±4.8. (рис 12) Таким образом наблюдаемое уменьшение квантового состава при увеличении температуры может быть связано с уменьшением пресинаптического уровня кальция, о чем говорит уменьшении кальциевого транзиента при увеличении температуры окружающей среды.
Рис. 12 Изменение кальциевого транзиента пресинаптической терминали двигательного нерва LAL мыши при увеличении температуры от контроля 20![]()
![]() С до 30
С до 30![]()
![]() С с шагом в 5
С с шагом в 5![]()
![]() С. Значения кальциевого транзиента на графике выражены в процентах от контроля. Различия достоверны, Р<0.05. Метод подсчета достоверности описан в методике выше. (n=5)
С. Значения кальциевого транзиента на графике выражены в процентах от контроля. Различия достоверны, Р<0.05. Метод подсчета достоверности описан в методике выше. (n=5)
3.4 Обсуждение
В ходе проделанной работы нами разработана методика загрузки кальциевого красителя в нервную терминаль теплокровных через культю нерва. Разработанный нами метод загрузки кальциевого флуоресцентного красителя через культю нерва имеет ряд преимуществ, перед другими методами загрузки флуоресцентных кальциевых индикаторов в живые клетки.
При работе на синапсах ЦНС в культуре нейронов или на срезах в основном применяют методики загрузки через пэтч-пипетку или используют формы красителей, проникающие сквозь клеточные мембраны – АМ-формы. Использование пэтч-пипеток при работе на нервно-мышечном препарате невозможно из-за его морфологического строения: малых размеров пресинаптической клетки, наличия соединительной ткани и т. д. Хотя применение этой методики допускается при работе на нервно-мышечных синапсах, выращенных в культурах клеток. Метод микроинъекций красителя в клетки был опробован нами, но после процедуры микроинъекции процент выживаемости препаратов был очень низок. При использовании формы красителей, проникающие сквозь клеточные мембраны – АМ-формы, трудно контролировать проникновение красителя в клеточные органеллы, хотя эта проблема может быть частично решена за счет выбора температурного режима инкубации препарата с красителем. Так же сложно дифференцировать сигналы от пресинаптической и мышечной клеток.
В методе загрузки через культю нерва краситель проникает только в цитозоль пресинаптической клетки, что исключает ошибки при регистрации сигналов, связанные с влиянием кальциевых сигналов от постсинаптической клетки или от внутриклеточных органелл, возникающие при других способах загрузки. После загрузки, проведенной данным способом, нервные окончания, находящиеся в проксимальной части препарата (ближе к культе нерва), равномерно окрашиваются красителем. Это позволяет проводить регистрацию флуоресцентных кальциевых сигналов от одного или нескольких нервных окончаний теплокровных
Согласно данным литературы изменение температуры окружающей среды может влиять на кальциевый транзиент и квантовый состав вызванных потенциалов концевой пластинки через изменение динамики работы кальциевых каналов [Iftinca et al., 2006]. Согласно этому исследованию, свойства кальциевых каналов и их функциональные роли могут меняться при приближении к физиологическим температурам. Исследования проводились при температурах 21 оС и 37 оС на разных изоформах кальциевых каналов теплокровных(крысы и человека).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
