

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
По-видимому, количество циклов клеточных делений в ходе онтогенеза генетически предопределено. Вместе с тем известна мутация, изменяющая размеры организма за счет одного дополнительного клеточного деления. Это мутация gt (giant), описанная у Drosophila melanogaster. Она наследуется по рецессивному сцепленному с полом типу. У мутантов gt развитие протекает нормально на протяжении всего эмбрионального периода. Однако в тот момент, когда нормальные особи окукливаются и начинают метаморфоз, особи gt продолжают оставаться в личиночном состоянии еще дополнительно 2—5 сут. За это время у них происходит одно, а может быть, и два дополнительных деления в имагинальных дисках, от количества клеток которых зависит размер будущей взрослой особи. Затем мутанты образуют куколку вдвое крупнее обычной. После метаморфоза несколько удлиненной по времени стадии куколки на свет появляется морфологически нормальная взрослая особь удвоенного размера.
У мышей описан ряд мутаций, обусловливающих снижение пролиферативной активности и следующие за этим фенотипические эффекты. К ним относят, например, мутацию or (ocular retardation), затрагивающую сетчатку глаза начиная с 10-х суток эмбрионального развития и приводящую к микрофтальмии (уменьшению размеров глазных яблок), и мутацию tgia, затрагивающую центральную нервную систему с 5—6-х суток после рождения и приводящую к отставанию роста и атрофии некоторых внутренних органов.
Таким образом, деление клеток является чрезвычайно важным процессом в онтогенетическом развитии. Оно протекает с разной интенсивностью в разное время и в разных местах, носит клональный характер и подвержено генетическому контролю. Все это характеризует клеточное деление как сложнейшую функцию целостного организма, подчиняющегося регулирующим влияниям на различных уровнях: генетическом, тканевом, онтогенетическом.
8.2.2. Миграция клеток
Миграции клеток, или клеточные перемещения, наряду с другими клеточными процессами имеют очень большое значение, начиная с процесса гаструляции и далее, в процессах морфогенеза. Клетки мезенхимного типа мигрируют одиночно и группами, а клетки эпителиев обычно согласованно, пластом. Мезенхима — это скопление веретеновидных или звездчатых клеток, погруженных в межклеточный матрикс. Эпителий — группы клеток, плотно прилежащих друг к другу боковыми стенками и имеющих апикальную и базальную поверхности. Как мезенхима, так и эпителии могут быть образованы из любого из трех зародышевых листков. Клетки мезенхимного типа наиболее подвижны, так как не образуют между собой стойких контактов.
Наиболее яркий пример миграции мезенхимных клеток связан с нервным гребнем. При смыкании нервной трубки клетки нервных валиков выходят из ее состава и располагаются между ее дорсальной частью и эктодермой (см. рис. 7.9).
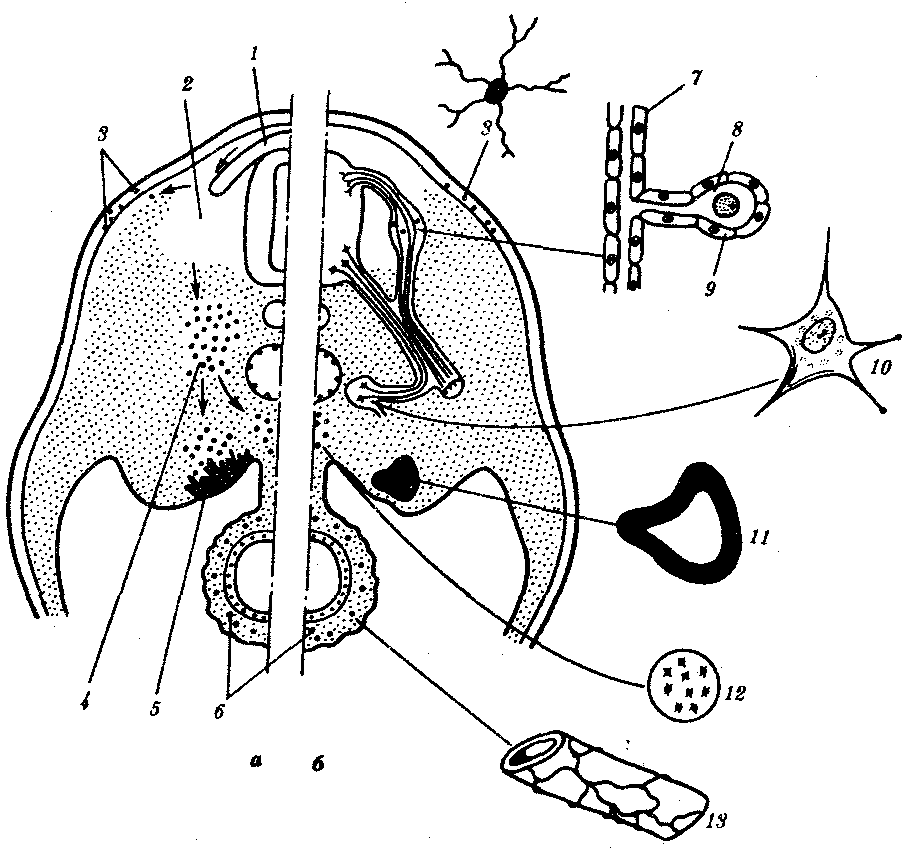
Рис. 8.1. Пути миграции клеток нервного гребня:
а—поперечный срез зародыша, б—производные клеток нервного гребня у взрослого организма; 1—нервный гребень, 2—узел спинного корешка, 3—пигментные клетки, 4—симпатический узел, 5—развивающийся надпочечник, 6—нервное сплетение в стенке кишки, 7— клетка шванновской оболочки, 8—униполярный чувствительный нейрон, 9—клетка-спутник, 10—мультиполярный нейрон симпатического узла, 11—хромаффинная клетка в мозговом веществе надпочечника, 12—превертебральное сплетение, 13— парасимпатическое сплетение в кишке; стрелками показано направление миграции клеток нервного гребня
Затем они мигрируют в разных направлениях, проявляя очень широкие формообразовательные потенции. Группа клеток нервного гребня в туловищной части зародыша мигрирует в эктодерму и там превращается в первичные пигментные клетки — меланоциты. Другие, двигаясь в центральном направлении, образуют нейроны спинальных ганглиев, еще дальше — ганглиев симпатической и парасимпатической систем. Третьи — превращаются в клетки шванновских оболочек нервов, четвертые — в хромаффинные клетки мозгового вещества надпочечников. Схематическое изображение этих миграций показано на рис. 8.1. Вообще клетки нервного гребня туловищного отдела дифференцируются в зависимости от того, куда попадут.
Клетки нервного гребня в головной части зародыша мигрируют в сторону лица, превращаясь в хрящевые, мышечные, соединительнотканные. Они строят хрящи висцерального скелета, мышцы кожи, соединительную ткань лица, языка и нижней челюсти, входят в состав аденогипофиза, паращитовидных желез и мякоти зуба. Если клетки головного отдела нервного гребня пересадить в туловищную область, то они все равно дифференцируются затем в хрящевые, мышечные и соединительнотканные. Еще одним убедительным примером выраженных миграций является перемещение первичных половых клеток из желточной энтодермы в зачаток половой железы.
Нарушение, миграции клеток в ходе эмбриогенеза приводит к недоразвитию органов или к их гетеротопиям, изменениям нормальной локализации. То и другое представляет собой врожденные пороки развития. Примеры пороков развития, связанных с нарушениями миграции клеток, известны, в частности, в отношении конечного мозга. Если нарушается миграция нейробластов, то возникают островки серого вещества в белом веществе, при этом клетки утрачивают способность к дифференцировке. Более выраженные изменения миграции приводят к микрогирии и полигирии (большое число мелких и аномально расположенных извилин больших полушарий), либо, наоборот, к макрогирии (утолщение основных извилин), либо к агирии (гладкий мозг, отсутствие извилин и борозд больших полушарий). Все эти изменения сопровождаются нарушением цитоархитектоники и послойного строения коры, гетеротопиями нервных клеток в белом веществе. Подобные пороки развития отмечены и в мозжечке.
Существуют гипотезы о дистантных воздействиях на клетки на основе хемотаксиса и о контактных воздействиях. Мезенхимные клетки способны к амебоидным движениям. Движение их по типу хемотаксиса показано для некоторых видов специализированных клеток (гоноциты, сперматозоиды, некоторые клетки крови). Для эмбриональных клеток многоклеточных животных достоверных случаев хемотаксиса не обнаружено. Контактные взаимодействия более распространены. Они представляют собой взаимодействие клеток со структурированным субстратом. Клетки ощущают микроструктуру субстрата и движутся вдоль волокон, как это происходит при помещении фибробластов на каплю кровяной плазмы, натянутой в виде пленки между сторонами стеклянного треугольника. На неструктурированной подложке, например на ненатянутой пленке, фибробласты перемещаются беспорядочно.
Хорошо известно также, что нервные волокна проходят часто длинный путь по различным тканям от нервного центра к рецептору или эффектору и «узнают» их. Скорее всего нервные окончания растут по микроструктурам субстрата, как они делают это и в культуре тканей. Субстратом для движения может быть и соседняя клетка, если оболочка ее натянута. А так как поверхность направленно движущейся клетки сама вытягивается, то и она может служить субстратом для движения и поляризации следующей за ней клетки.

Рис. 8.2. Перемещение клеточных пластов.
А —путем выпячивания на примере образования глазного пузырька; Б—путем впячивания на примере образования слухового пузырька:
а — стенка переднего мозга, б —местное ускорение роста, в — выпячивание, г — глазной пузырек, д—ж —углубление ямки, з —отшнуровывание пузырька
Согласованные перемещения пластов эпителиальных клеток также изучаются. К ним относятся изгибы клеточных пластов путем выпячивания или впячивания, отшнуровка, образование утолщений — плакод (рис. 8.2).
Таким, образом, несомненно, что для миграции клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. И то, и другое генетически детерминировано, так что и сама миграция клеток находится под генетическим контролем, с одной стороны, и влияниями окружающих клеток и тканей — с другой.
8.2.3. Сортировка клеток
В процессе эмбриогенеза клетки не только активно перемешаются, но и «узнают» друг друга, т. е. образуют скопления и пласты только с определенными клетками. Значительные координированные перемещения клеток характерны для периода гаструляции. Смысл этих перемещений заключается в образовании обособленных друг от друга зародышевых листков с совершенно определенным взаимным расположением. Клетки как бы сортируются в зависимости от свойств, т. е. избирательно.
Замечено, что необходимым условием сортировки являются степень подвижности клеток иособенности их мембран. В поздней бластуле амфибий, например, клетки будущей эктодермы обладают тенденцией слипаться друг с другом и распространяться в виде сплошного слоя над мезодермой и энтодермой. Эта тенденция проявляется и в культуре тканей. Клетки мезодермы имеют тенденцию впячиваться в любой находящийся поблизости комок клеток, а клетки энтодермы относительно неподвижны.
Эти особенности трудно объяснить, если наблюдать только за нормальным течением развития. Впервые на различия в подвижности и способности к слипанию (адгезии) между клетками, относящимися к разным зародышевым листкам, обратил внимание Г. Гольтфретер в 30-х гг. XX столетия. Затем производили модификации его опытов с самыми разными клетками. Смысл этих опытов состоит в том, что зародыши тритонов или других животных на стадии гаструлы диссо-циируют с помощью фермента трипсина, который разрушает материал, соединяющий клетки друг с другом. Диссоциированные (дезагрегированные) клетки тщательно перемешивают и затем создают такие условия, чтобы клетки могли свободно перемещаться и воссоединяться друг с другом. Сначала клетки представляют собой беспорядочную смесь, затем клетки эктодермы, мезодермы и энтодермы разделяются (сегрегируют) и собираются в отдельные группы, каждая из которых представляет собой клеточный агрегат из однородных клеток. Образуются снова зародышевые листки, располагающиеся иногда даже в обычном для них порядке (рис. 8.3).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 |
