

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Для количественного определения нуклеиновых кислот по их углеводным компонентам используют цветные реакции дезоксирибозы с фениламином (для ДНК) и рибозы с орцином (для РНК).
Работа 3.2. Количественное определение ДНК
по методу Дише
Принцип метода. При нагревании в присутствии концентрированных кислот 2-дезоксирибоза (углеводный компонент ДНК) образует фурфуриловый спирт и другие продукты, которые затем концентрируются с ароматическим аминами с образованием комплексов синего цвета. Чаще всего при определении этим методом используют дифениламин:
15
NH |


Реактив Дише: 1 г дифениламина растворяют в 100 мл ледяной уксусной кислоты и добавляют 2,75 мл концентрированной серной кислоты. Реактив готовят перед определением.
Ход работы: Готовят две пробирки. В опытную пробирку отмеривают 1 мл водного раствора ДНК и 2 мл дифениламинового раствора. Обе пробы помещают в кипящую водяную баню на 10 мин. Затем пробы охлаждают и измеряют интенсивность окраски на фотоэлектроколориметре (ФЭК) против контроля (светофильтр красный). Зная оптическую плотность опытной пробы, по калибровочному графику находят в ней ДНК.
Построение калибровочного графика. В три пробирки наливают по 1 мл раствора ДНК с различной концентрацией (50, 100, 200 мкг/мл) и по 2 мл дифениламиного реактива. Помещают пробирки на 10 мин в кипящую водяную баню, а затем измеряют оптическую плотность каждого из растворов. Калибровочный график строят, откладывая по оси абцисс концентрации использованных растворов ДНК, а по сои ординат – соотвествующие им значения оптической плотности.
Оформление работы. Записать принцип метода, результаты колориметрии, построить график.
Работа 3.3. Количественное определение РНК
по методу Майбаум
Принцип метода. Рибоза при конденсации с орцином образует окрашенные в зеленый цвет продукты. Орцин представляет собой метилированное производное резорцина:
16
CH3 НО |
 НО
НОРеактив. 50 мг FeCl3 в 250 мл концентрированной HCl. Непосредственно перед определением добавляют орцин из расчета 4,76 мг на 1 мл раствора. Хлорное железо служит катализатором реакции.
Ход работы. В опытную пробирку отмеривают 1 мл раствора РНК и 1 мл орцинового реактива. В контрольную пробирку отмеривают 1 мл дистилированной воды и 1 орцинового реактива. Обе пробирки помещают в кипящую водяную баню на 20 мин. После охлаждения измеряют интенсивность окраски на ФЭКе (красный светофильтр) против контроля. Измерив оптическую плотность опытной пробы, по калибровочному графику находят концентрацию РНК в данной пробе.
Построение калибровочного графика. В три пробирки наливают по 1 мл раствора РНК с известной концентрацией (50, 100, 200 мкг/мл) и по 1 мл орнитинового реактива. Пробы помещают в кипящую водяную баню на 20 мин и после охлаждения измеряют оптическую плотность каждого из растворов. Затем строят калибровочный график, откладывая по оси абцисс концентрацию РНК, а по оси ординат соответствующее им значение оптической плотности.
Оформление работы. Записывают в рабочую тетрадь принцип метода, результаты колориметрии и калибровочный график.
Определение нуклеиновых кислот по содержанию фосфора
Определение по фосфору считается наиболее надежным методом оценки количества нуклеиновых кислот, так как его содержание в наименьшей степени зависит от нуклеотидного со
17
става: для ДНК оно составляет 9,8 – 10,1%, для РНК – 9.1 – 9,6%. При определении фосфора нуклеиновых кислот в тканях необходимо предварительно удалить свободные нуклеотиды, неорганический фосфат, а также все фосфоросодержащие соединения ненуклеотидной природы, в частности, липиды.
Работа 3.4. Количественное определение ДНК
по содержанию фосфора
Ход работы. В колбу для сжигания приливают 1 мл исследуемого раствора ДНК и добавляют 1 мл смеси для сжигания (концентрированная H2SO4 + HclO4). Нагревают на плитке до образования раствора. Затем колбы охлаждают, добавляют 10 мл воды и нейтрализуют по фенолфталеину (добавляют его 1 – 2 капли в колбу) 10% раствором NaOH до слаборозовой окраски. Содержимое колбы переливают в мерную колбу на 25 мл и доводят водой до метки.
Определение фосфора. В мерную колбу на 10 мл налить 2 мл минерализата, 0,6 мл амидола и 1,6 мл молибденовокислого аммония и довести водой до метки. Оставить на 30 минут для развития окраски. Колориметрировать при 830 нм.
Содержание фосфора определяют по калибровочной кривой и рассчитывают по формуле:
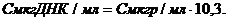
Домашнее задание: подготовиться к семинару по теме " Метилирование ДНК ".
Занятие VI
Теоретическая часть
Семинарское занятие по теме "Метилирование ДНК".
18
1. Метилирование цитозина в ДНК эукариот.
2. История открытия явления рестрикции-модификации у бактерий.
3. Системы рестрикции-модификации I, II и III типов.
4. Роль метилирования в репарации ДНК.
Практическая часть
Решение задач
1. Определить размер фрагментов, которые образуются при гидролизе ДНК фага-ЛЯМБДА рестриктазой KpnI, имеющей сайты рестрикции в положении 17 058 и 18 561. ДНК этого фага имеет линейную молекулу длиной 48 502 п. н.
2. Плазмида pBR322 размером 4363 п. н. имеет 3 сайта рестрикции BglI в точках 936, 1 170, 3 489. Определить размер фрагментов, которые получатся при гидролизе молекулы ДНК данной рестриктазой.
3. Рестриктазы BamHI и PstI разрезают узнаваемые ими последовательности, как показано на рис. 1.
5` 3`
-- Г Г А Т Ц Ц -- BamHI -- Г Г А Т Ц Ц --
![]() -- Ц Ц Т А Г Г -- -- Ц Ц Т А Г Г --
-- Ц Ц Т А Г Г -- -- Ц Ц Т А Г Г --
![]() 3` 5`
3` 5`
5` 3`
-- Ц Т Г Ц А Г -- PstI -- Ц Т Г Ц А Г --
![]() -- Г А Ц Г Т Ц -- -- Г А Ц Г Т Ц --
-- Г А Ц Г Т Ц -- -- Г А Ц Г Т Ц --
![]() 3` 5`
3` 5`
Рис. 1. Расщепление рестриктазами BamHI и PstI узнаваемых ими последовательностей. Показаны нуклеотиды, образующие сайты узнавания.
19
А. Укажите, где находятся 5`- и 3`-концы разрезаемых молекул ДНК.
Б. Как будут модифицироваться эти концы, если инкубировать разрезанные молекулы ДНК с ДНК-полимеразой в присутствии всех четырех дезоксинуклеозидтрифосфатов?
В. Могут ли концы, появившиеся в результате разрезания ферментом BamHI, соединиться вновь при инкубации с ДНК-лигазой фага Т4 после того, как была проведена реакция, о которой шла речь в пункте Б?
Можно ли соединить вместе концы ДНК, образовавшиеся при обработке PstI?
(Лигаза ДНК фага Т4 соединяет вместе "тупые" концы так же, как и "липкие").
Г. Будет ли регенерировать узнаваемый BamHI сайт, о котором шла речь в пункте В, при соединении концов? Будет ли регенерировать сайт, узнаваемый PstI?
4. Фермент рестрикции Sau3A узнает последовательность – ГАТЦ— и расщепляет цепь ДНК на 5`-стороне (слева) от Г (поскольку верхняя и нижняя цепи большинства сайтов рестрикции читаются одинаково в направлении от 5`- к 3`-концу, только одна цепь этого сайта может быть показана для иллюстрации). Одноцепочечные концы, возникающие при расщеплении рестриктазой Sau3A, идентичны концам, возникающим при расщеплении рестриктазой BamHI (рис. 1), что позволяет им соединяться при инкубации с ДНК-лигазой.
А. Какая доля сайтов, узнаваемых BamHI, может быть разрезана рестриктазой Sau3A? Какая доля сайтов, узнаваемых Sau3A, может быть разрезана рестриктазой BamHI?
Б. Если два конца, образовавшиеся при расщеплении BamHI, сшиваются вместе, то образующийся сайт может быть снова расщеплен BamHI. То же самое справедливо для двух концов, образующихся при расщеплении Sau3A. Предположите, однако, что вы сшиваете конец, образовавшийся при обработке Sau3A, с концом, образовавшимся при обработке BamHI. Может ли гибридный сайт разрезаться рестриктазой Sau3A, рестриктазой BamHI? 20
В. Каков будет средний размер фрагментов ДНК, образующихся при обработке хромосомной ДНК рестриктазой Sau3A, рестриктазой BamHI?
5. Постройте рестрикционную карту по результатам гель-электрофореза фрагментов ДНК, полученных расщеплением молекулы указанными ферментами. Очищенную ДНК гидролизовали рестриктазами EcoRI и BamHI раздельно, а затем их смесью, проводили гель-электрофорез и визуализировали продукты окрашиванием бромистым этидием. Числа слева от горизонтальных полос – длина фрагментов в парах оснований.
850 500 300 | BamHI 950 600 100 | EcoRI + BamHI 600 400 300 250 100 |


Домашнее задание: подготовиться к семинару "Транскрипция".

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
