

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В. Если площадь мембран всех других компартментов (лизосом. эндосом, внутренней ядерной мембраны, секреторных везикул), которые получают мембранные белки из ЭР, в сумме равны площади плазматической мембраны, то какова доля в ЭР тех мембранных белков этой клетки, которые постоянно присутствуют в мембране ЭР?
2.Время жизни молекулы белка обычно соответствует той функции, которую он выполняет in vivo; например, структурные белки относятся к долгоживущим, тогда как регуляторные белки - это, как правило, короткоживущие молекулы. Время жизни белка в эукариотической клетке может сильно зависеть от его N-концевой аминокислоты. В опытах, с помощью которых было показано влияние N-концевых аминокислот на стабильность белка, использовали гибридный белок, состоявший из убиквитина, ковалентно сшитого с β-галактозидазой. Исследователи, обнаружившие это явление, изучили разнообразные плазмиды, кодирующие различные варианты гибридного белка. Три такие плазмиды схематически представлены на рис. 1. Когда эти плазмиды вводили в дрожжевые клетки, последние синтезировали гибридные белки, но убиквитин отщеплялся (неидентифицированным ферментом) точно по месту его соединения с β - галактозидазой. В результате возникали молекулы β-галактозидаз с разными N-концами ( рис. 1).
28
|
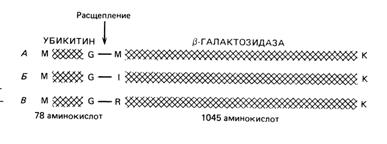
Рис. 1. Гибридные белки, кодируемые тремя дрожжевыми плазмидами. При экспрессии в клетках дрожжей эти белки расщепляются по пептидным связям, указанным стрелкой. Молекулы β-галактозидаз, освобождающиеся при этом, различаются только по N-концам.
Чтобы оценить период полужизни этих β-галактозидаз, дрожжи выращивали на протяжении нескольких поколений в присутствии радиоактивной аминокислоты. Затем синтез белков блокировали с помощью соответствующего ингибитора. Для определения скорости деградации β-галактозидазы из культуры отбирали пробы в разные моменты времени, проводили очистку β - галактозидазы с помощью специфических антител и измеряли количество радиоактивной β-галактозидазы после гель-электрофореза с детергентом – додецилсульфатом натрия (ДСН). Результаты электрофореза для пробы, отобранной через 5 мин после добавления ингибитора, представлены на рис. 2,А. Динамика деградации за весь период отбора проб показана на рис. 2,Б.
А. С помощью метода рекомбинантной ДНК можно было бы создать ряд плазмид с измененным первым кодоном гена β-галактозидазы. Как вы думаете, почему не был использован этот более прямой путь?
Б. Оцените период полужизни трех различных видов β-галактозидаз (время, за которое деградирует половина материала).
29
|
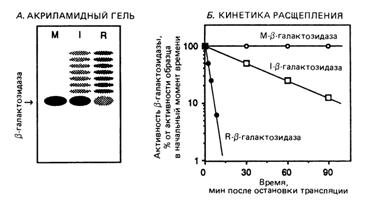
Рис. 2. Результаты опытов с тремя различными гибридными белками. А. Электрофоретическое выделение радиоактивной β-галактозидазы, полученной путем осаждения с помощью антител, N-концевые аминокислоты указаны над каждым столбиком. Б. Динамика деградации β-галактозидазы после завершения синтеза белка. Уровень β-галактозидазы выражается в процентах от начального количества, имевшегося сразу после остановки синтеза белка.
3.Когда вы узнали о работе, описанной в предыдущей задаче, вас заинтересовали два аспекта представленных там данных. Во-первых, действительно ли убиквитин удалялся с N-конца? Во-вторых, чему соответствуют полосы, расположенные над полосами β-галактозидазы на электрофореграмме (рис. 2, А)? Любые фрагменты β-галактозидазы, образующиеся при ее деградации, должны находиться на электрофореграмме ниже полосы β-галактозидазы.
Чтобы проверить, действительно ли β-галактозидаза высвобождается в результате отщепления убиквитина от образовавшегося гибридного белка, вы выделяете с помощью антител нерадиоактивную β-галактозидазу из клеток, содержащих те же три плазмиды. Затем вы подвергаете эти образцы гель-электрофорезу с ДСН, переносите пластины геля на фильтровальную бумагу и проводите реакцию взаимодействия с радиоактивными антителами против убиквитина. Как видно из рис. 3,
30
антитела к убиквитину не взаимодействовали с материалом, соответствующим полосе β-галактозидазы. Однако эти антитела взаимодействовали со всеми полосами, расположенными над β-галактозидазой. Расположение этих полос было таким же, как и расположение полос на рис. 2, А.
А. Показывают ли эти опыты, что убиквитин отщепляется от N-конца гибридного белка?
Б. Почему в случаях, когда на N-конце находились остатки изолейцина (I) или аргинина (R), убиквитин присутствовал в полосах, расположенных выше полосы (β-галактозидазы, а в тех случаях, когда на N-конце был остаток метионина (М), убикитин не обнаруживался?
4.Орнитиндекарбоксилаза (ОДКаза) катализирует первую реакцию в биосинтезе полиаминов, которые связываются с сахарофосфатным остовом молекул ДНК и РНК и играют существенную роль в процессах роста клеток. Активность ОДКазы регулируется обычно в соответствии с физиологическим состоянием клетки.
|
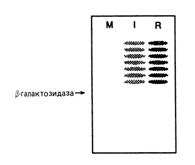
Рис. 3. Электрофорез немеченой β-галактозидазы и последующее взаимодействие с мечеными антителами к убиквитину. Сокращенные обозначения N-концевых аминокислот указаны над столбиками. Положение нативной β-галактозидазы указано стрелкой.
Представьте, что вы выделили мутантную линию клеток печени с гораздо более высокой, чем в норме, активностью ОДКазы. Ваши предварительные опыты позволяют предположить,
31
что нормальные и мутантные клетки несут одинаковое число генов для ОДКазы, что эти гены транскрибируются с одинаковой скоростью и что нет явных различий в структурных и ферментативных свойствах ОДКаз из нормальных и мутантных клеток. Однако когда вы добавляете ингибитор синтеза белка, циклогексимид, и измеряете затем ферментативную активность в разные моменты времени, то обнаруживаете, что ОДКаза из мутантных клеток имеет период полужизни 12 ч вместо 14 мин, как у ОДКазы из обычных клеток. Период полужизни других белков у мутантной линии не изменялся.
А. Если ОДКаза образуется с той же скоростью, что и в нормальных клетках, то почему в мутантных клетках активность этого фермента гораздо выше?
Б. Каким образом можно было бы определить, чем обусловлена нестабильность ОДКазы в нормальных клетках: разрушением белка или инактивацией его в результате модификации (например, фосфорилирования, метилирования или ацетилирования)?
В. Предполагая, что нестабильность ОДКазы в нормальных клетках объясняется деструкцией белка, предложите объяснение стабильности ОДКазы в клетках мутантной линии.
2. Лабораторная работа № 7
Методы количественного определения белков
Для количественного определения белков применяют физические, химические и биологические методы.
Из физических методов простейшим кажется взвешивание чистого белка. Однако белки очень гигроскопичны, и полностью удалить из их состава воду столь трудно, что этот способ количественного определения белков применяют редко. Кроме того, выделить весь белок из препарата практически невозможно.
32
Наибольшее распространение из физических методов количественного определения белков получили три: рефрактометрический (по показателю преломления белковых растворов), спектрофотометрический (по поглощению в ультрафиолетовой области спектра) и полярографический (по кривым, показывающим зависимость между силой тока и напряжением, приложенным к системе, содержащей белок). Пикнометрический метод (по плотности белковых растворов) употребляется редко.
Химические методы количественного определения белков разнообразны. Наиболее простым химическим методом определения белка является количественное определение общего или белкового (после осаждения белка и отделения его от растворимых азотсодержащих веществ) азота. Умножая величину процентного содержания общего азота на коэффициент 6,25 (среднее содержание азота в белках – 16 %, отсюда 100 : 16 = 6,25), получают данные о содержании сырого протеина. Проделывая ту же операцию с величиной, характеризующей содержание белкового азота, получают данные о количестве белка. Эти способы условны, ибо не дают абсолютных результатов.
На том же принципе основаны два других метода химического определения белков: по содержанию металла и по содержанию той или иной аминокислоты. Например, в гемоглобине содержится 0,34% железа. Если в изучаемом на содержание гемоглобина препарате нет других железосодержащих соединений, то определение в препарате железа дает возможность рассчитать содержание гемоглобина. Аналогично рассуждают, если в составе препарата определено содержание какой-либо аминокислоты, доля которой в белке хорошо известна. Оба перечисленных метода применяются лишь в отдельных случаях.
Самым распространенным химическим методом количественного определения белков является колориметрический метод. Он основан на измерении интенсивности цветных реакций, развивающихся при взаимодействии белков с тем или иным специфическим реагентом. Чтобы рассчитать концентрацию белка, в этом случае строят калибровочный график.
33
Биологические методы количественного определения белков применимы лишь к белкам, обладающим ферментативной и гормональной активностью. Измеряя степень биологической активности препарата, можно составить представление о содержании в нем белка, обладающего данной активностью. Этот метод тоже не дает абсолютных результатов.
Количественное определение белка по биуретовой реакции
Биуретовая реакция белков не отличается высокой чувствительностью. Поэтому она применяется в тех случаях, когда содержание белка в исследуемом образце достаточно велико (не ниже нескольких миллиграммов на миллилитр).
Оборудование, реактивы. Фотоэлектроколориметр; пробирки стеклянные химические; пипетки градуированные на 1,2 и 10 мл; гидроксид натрия (10%-ный), не содержащий примеси карбонатов (готовится разведением 75%-ного раствора гидроксида натрия, в котором карбонаты нерастворимы, прокипяченной дистиллированной водой, свободной от СО2); хлорид натрия (1%-ный); сульфат меди; сегнетова соль; сыворотка крови; гемолимфа насекомых; экстракты тканевых белков.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
