

Энергия электрохимического протонного градиента используется для синтеза АТР и транспорта метаболитов и неорганических ионов в матрикс [9]
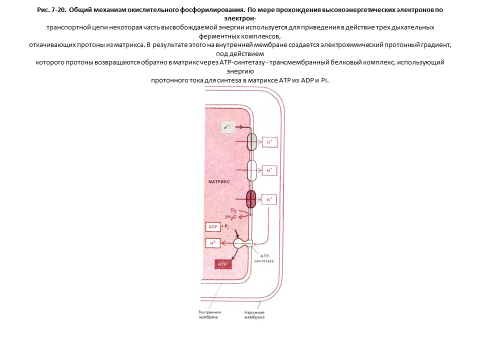
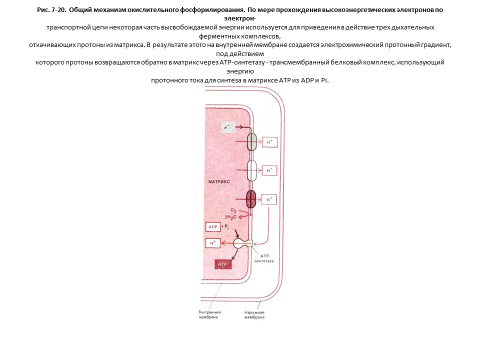
Внутренняя мембрана митохондрий отличается необычно высоким содержанием белка - в ней по весу примерно 70% белка и 30%фосфолипидов. Многие из этих белков входят в состав электронтранспортной цепи, поддерживающей протонный градиент на мембране. Другой важный компонент - фермент АТР-синтетаза, катализирующий синтез АТР. Это большой белковый комплекс, через который протоны перетекают обратно в матрикс по электрохимическому градиенту. Подобно турбине, АТР-синтетаза преобразует одну форму энергии в другую, синтезируя АТР из ADP и Рi в митохондриальном матриксе в ходе реакции, сопряженной с током протонов в матрикс (рис. 7-20).
Но синтез АТР - это не единственный процесс, идущий за счет энергии электрохимического градиента. В матриксе, где находятся ферменты, участвующие в цикле лимонной кислоты и других метаболических реакциях, необходимо поддерживать высокие концентрации различных субстратов; в частности, для АТР-синтетазы требуются ADP и фосфат. Поэтому через внутреннюю мембрану должны транспортироваться разнообразные несущие заряд субстраты. Это достигается с помощью различных белков-переносчиков, встроенных в мембрану (см. разд. 6.4.4), многие из которых активно перекачивают определенные молекулы против их электрохимических градиентов, т. е. осуществляют процесс, требующий затраты энергии. Для большей части метаболитов источником этой энергии служит сопряжение с перемещением каких-то других молекул «вниз» по их электрохимическому градиенту (см. разд. 6.4.9). Например, в транспорте ADP участвует система антипорта ADP-ATP: при переходе каждой молекулы ADP в матрикс из него выходит по своему электрохимическому градиенту одна молекула АТР. В то же время система симпорта сопрягает переход фосфата внутрь митохондрии с направленным туда же потоком Н+: протоны входят в матрикс по своему градиенту и при этом «тащат» за собой фосфат. Подобным образом переносится в матрикс и пируват (рис. 7-21).
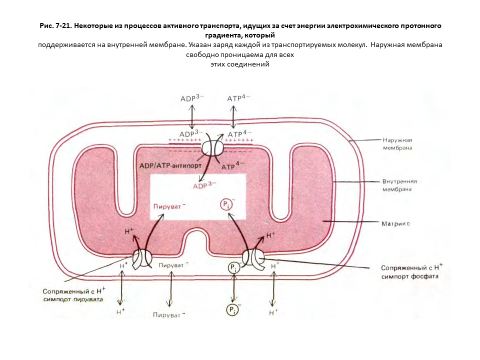
Энергия электрохимического протонного градиента используется также для переноса в матрикс ионов Са2+ , которые, по-видимому, играют важную роль в регуляции активности некоторых митохондриальных ферментов; большое значение может иметь и поглощение митохондриями этих ионов для удаления их из цитозоля, когда концентрация Са2+ в последнем становится высокой (см. разд. 12.3.7).
Чем больше энергии электрохимического градиента затрачивается на перенос молекул и ионов в митохондрию, тем меньше остается для синтеза АТР. Например, если изолированные митохондрии поместить в среду с высоким содержанием Са2+ , то они полностью прекратят синтез АТР; вся энергия градиента будет расходоваться на транспорт Са2+ в матрикс. В некоторых специализированных клетках электрохимический протонный градиент «шунтируется» таким образом, что митохондрии вместо синтеза АТР образуют тепло (см. разд. 7.2.12). Очевидно, клетки способны регулировать использование энергии электрохимического протонного градиента и направлять ее на те процессы, которые наиболее важны в данный момент.
Быстрое превращение ADP в АТР в митохондриях позволяет поддерживать высокое отношение концентраций ATP/ADP в клетках [10]
С помощью особого белка, встроенного во внутреннюю мембрану, ADP транспортируется в матрикс в обмен на АТР по принципу антипорта. В результате молекулы ADP, высвобождаемые при гидролизе АТР в цитоплазме, быстро поступают в митохондрию для «перезарядки», в то время как молекулы АТР, образующиеся в матриксе в процессе окислительного фосфорилирования, тоже быстро выходят в цитозму, где они нужны. В организме человека молекулы АТР за сутки оборачиваются несколько тысяч раз, что позволяет поддерживать в клетке концентрацию АТР, более чем в 10 раз превышающую концентрацию ADP.
Ранее я говорил, что ферменты клетки превращают субстраты зачастую осуществляя энергетически невыгодные реакции путем сопряжения их с энергетически выгодным гидролизом АТР (см. рис. 2-27). 
Таким образом, пул АТР обеспечивает внутриклеточные процессы энергией подобно аккумулятору, приводящему в действие электромотор: если митохондрии прекратят свою активность, то клеточная «аккумуляторная батарея» начнет разряжаться и наступит момент, когда энергетически невыгодные реакции уже не смогут осуществляться за счет гидролиза АТР.
На первый взгляд может показаться, что такого состояния не будет до тех пор, пока концентрация АТР не упадет до нуля. Фактически же это состояние наступает значительно раньше - при определенном уровне АТР, зависящем от концентраций ADP и Рi. Для того чтобы объяснить, почему так происходит, нужно обратиться к некоторым элементарным принципам термодинамики.
Клеточное дыхание необычайно эффективно
В процессе окислительного фосфорилирования каждая пара электронов NADH обеспечивает энергией образование примерно трех молекул АТР. Пара электронов FADH2, обладающая меньшей энергией, дает энергию для синтеза только двух молекул АТР. В среднем каждая молекула ацетил-СоА, поступающая в цикл лимонной кислоты, дает около 12 молекул АТР. Это означает, что при окислении одной молекулы глюкозы образуются 24 молекулы АТР, а при окислении одной молекулы пальмитата - жирной кислоты с 16 углеродными атомами - 96 молекул АТР.
Если учесть также экзотермические реакции, предшествующие образованию ацетил-СоА, окажется, что полное окисление одной молекулы глюкозы дает около 36 молекул АТР, тогда как при полном окислении пальмитата образуется примерно 129 молекул АТР. Это максимальные величины, так как фактически количество синтезируемого в митохондриях АТР зависит от того, какая доля энергии протонного градиента идет на синтез АТР, а не на другие процессы.
Если сравнить изменение свободной энергии при сгорании жиром и углеводов прямо до СО2 и Н2О с общим количеством энергии запасаемой в фосфатных связях АТР в процессах биологического окисления, окажется, что эффективность преобразования энергии окисления в энергию АТР часто превышает 50%. Это значительно выше эффективности большинства энерго-преобразующих устройств, созданных человеком.
Если бы клетка работала с эффективностью (к. п.д) электромотора или автомобильного двига%), то организму для поддержания жизни
требовалось бы намного больше пищи. Кроме того, поскольку вся неиспользованная энергия высвобождается в виде тепла, крупные организмы
нуждались бы в более эффективных способах отвода тепла в окружающую среду.
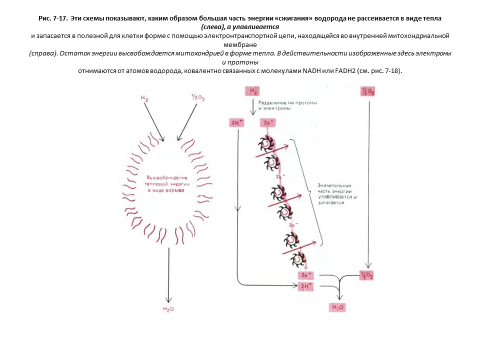
Изучая клеточное дыхание, студенты иногда удивляются, почему химические взаимопревращения в клетке идут таким сложным путем. Казалось бы, вполне можно обойтись без цикла лимонной кислоты и многих звеньев дыхательной цепи и окислять сахара до СО2 и Н2О более прямым способом. Но, для клетки подобный путь оказался бы катастрофическим. Огромное количество свободной энергии, высвобождаемое при окислении, может эффективно использоваться только мелкими порциями. В сложном процессе окисления участвует много промежуточных продуктов, каждый из которых лишь незначительно отличается от предыдущего. Благодаря этому высвобождаемая энергия дробится на меньшие количества, которые можно эффективно преобразовывать с помощью сопряженных реакций в высокоэнергетические связи молекул АТР и NADH (см. рис. 2-17).
ИТАК
Митохондрии осуществляют большую часть клеточных процессов окисления и производят почти весь АТР животной клетки.
Митохондриальный матрикс содержит множество разнообразных ферментов, в том числе ферменты, окисляющие пируват и жирные кислоты до ацетил-СоА, и ферменты, окисляющие этот ацетил-СоА до СОг в цикле. лимонной кислоты. В ходе этих реакций окисления образуютсябольшие количества NADH (и FADH2). Энергия, получаемая при соединении кислорода с переносимыми NADH и FADH2 реакционноспособными электронами, используется электронтранспортной цепью, находящейся во внутренней мембране митохондрии и называемой дыхательной цепью.
Дыхательная цепь «откачивает» протоны из матрикса, что приводит к созданию трансмембранного электрохимического протонного градиента, слагающегося из мембранного потенциала и разности рН. Энергия трансмембранного градиента в свою очередь используется для синтеза АТР и для активного транспорта необходимых метаболитов через внутреннюю митохондриальную мембрану. Сочетание этих реакций обеспечивает эффективный обмен ATP-ADP между митохондрией и цитозолем, что позволяет поддерживать в клетке высокий уровень АТР
АТР-синтетаза может действовать в обратном направлении - расщеплять АТР и перекачивать протоны [16]
Действие АТР-синтетазы обратимо: она способна использовать как энергию гидролиза АТР для перекачивания протонов через внутреннюю митохондриальную мембрану, так и энергию потока протонов по электрохимическому градиенту для синтеза АТР (рис. 7-26).
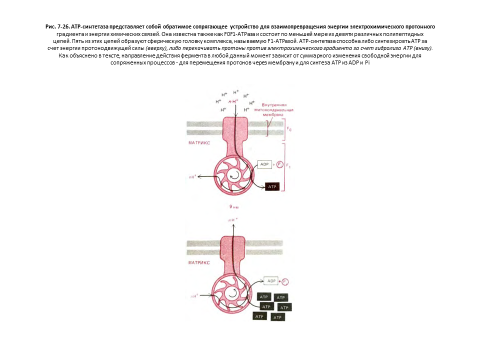
Таким образом, АТР-синтетаза - это обратимая сопрягающая система, которая осуществляет взаимопревращение энергии электрохимического протонного градиента и химических связей. Направление ее работы зависит от соотношения между амплитудой протонного градиента и локальной величиной ?G для гидролиза АТР.
АТР-синтетаза получила свое название в связи с тем, что в обычных условиях протонного градиента, обеспечиваемого дыхательной цепью (см. рис. 7-20), синтезирует большую часть всего АТР клетки. Для упрощения приводимых ниже расчетов мы будем предполагать, что при прохождении через АТР-синтетазу каждых трех протонов синтезируется одна молекула АТР.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
