

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Молекулярная биофизика. Биофизика трансформации энергии: митохондрии и хлоропласты
Митохондрии, обнаружены во всех эукариотических клетках, и свойственные только растениям пластиды (из которых наибольший интерес представляют хлоропласты) преобразуют энергию в формы, которые могут быть использованы для биоэнергетики клетки.
Специфическую функцию этих органелл отражает наиболее поразительная черта их морфологии - обилие внутренних мембран.
Мембраны выполняют в этих энергопреобразующих органеллах две ключевые функции.
Во-первых, они осуществляют процессы переноса электронов, в результате которых энергия реакций окисления (см. разд. 2.2.4) преобразуется в более полезные формы, главным образом в энергию АТР.
Во-вторых, мембраны образуют в органелле большие внутренние компартменты, в которых находятся ферменты, катализирующие другие внутриклеточные реакции.
Без митохондрий животная клетка могла бы получать АТР только за счет анаэробного гликолиза. Но в результате превращения глюкозы в пируват, происходящего при гликолизе (разд. 2.3.2), высвобождается лишь малая часть всей свободной энергии, которую можно получить при окислении сахаров. В митохондриях метаболизм сахаров (и жирных кислот) доводится до конца: пируват (как и жирные кислоты) окисляется молекулярным кислородом (О2) до СО2 и Н2О. Энергия, высвобождаемая при таком окислении, используется настолько эффективно, что на каждую молекулу окисляемой глюкозы образуется около 36 молекул АТР, в то время как при гликолизе на одну молекулу глюкозы приходится только 2 молекулы АТР.
Хлоропласты - это очень эффективные «машины» для выработки АТР, но источником энергии для них служит солнечный свет, а не сахара и жирные кислоты. Несмотря на такое фундаментальное различие, митохондрии и хлоропласты организованы сходно и синтезируют АТР одним и тем же способом.
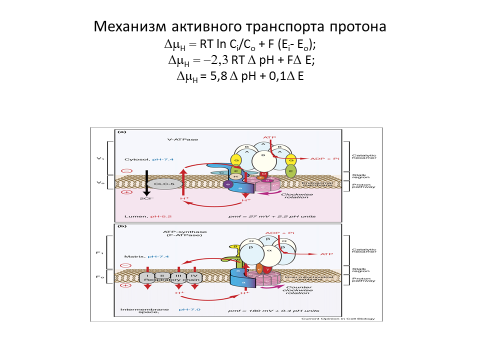
Общий путь, по которому митохондрии, хлоропласты и даже бактерии преобразуют энергию для биологических целей, основан на процессе, получившем название хемиосмотического сопряжения. Этот процесс начинается с того, что электроны, передаются от доноров по цепи из переносчиков электронов, встроенных в мембрану. При таком переносе по электронтранспортной цепи электроны, которые были извлечены либопри поглощении свет, либо - при окислении питательных веществ,
последовательно переходят на все более низкие энергетические уровни.
При этом, часть высвобождаемой энергии используется для перемещения протонов с одной стороны мембраны на другую, в результате чего на мембране создается электрохимический протонный градиент. За счет энергии этого градиента протекают реакции, катализируемые ферментами, встроенными в ту же мембрану (рис. 7-1). В митохондриях и хлоропластах большая часть энергии используется для превращения ADP и Рi в АТР, хотя некоторая ее доля расходуется на транспорт специфических метаболитов в органеллу и из нее. В отличие от этого у бактерий электрохимический градиент служит источником энергии, как и синтезируемый с его помощью АТР: благодаря энергии градиента осуществляются не только многие транспортные процессы, но и быстрое вращение бактериальных жгутиков, перемещающих клетку (разд. 12.5.4).
Как полагают, преобразующие энергию органеллы эукариот произошли от прокариотических клеток, которые были захвачены примитивными эукариотами и вступили с ними в симбиоз 1,5 млрд. лет назад. Этим можно объяснить, почему митохондрии и хлоропласты имеют свою собственную ДНК, кодирующую некоторые белки этих органелл. Однако к настоящему времени митохондрии и хлоропласты утратили большую часть собственного генома и стали полностью зависеть от белков, кодируемых ядерными генами, синтезируемых в цитозоле и лишь затем переносимых в органеллу. И наоборот, клетки-хозяева стали зависимы от этих органелл, дающих значительную часть АТР, необходимого для биосинтезов, активного транспорта ионов и растворенных веществ и двигательных функций, а также содержащих ряд ферментов, катализирующих некоторые реакции биосинтеза.
Митохондрии
Митохондрии занимают значительную часть цитоплазмы почти во всех эукариотических клетках. Хотя митохондрии настолько велики, что их можно увидеть в обычный световой микроскоп, и впервые были обнаружены еще в прошлом веке, все же реальная возможность разобраться в их функции появилась только после 1948 г., когда были разработаны методы выделения интактных митохондрий. По техническим причинам большинство биохимических исследований проводилось на митохондриях, выделенных из печени.
Митохондрии обычно представляют в виде жестких вытянутых, похожих на бактерии цилиндров диаметром от 0,5 до 1 мкм. Однако цейтраферная микрокиносъемка живых клеток позволяет увидеть, что митохондрии - необыкновенно подвижные и пластичные органеллы, которые постоянно изменяют свою форму (рис. 7-2) и даже сливаются друг с другом и затем вновь разделяются. Пути перемещения митохондрий в цитоплазме часто связаны с микротрубочками (рис. 7-3), что может определять характерную ориентацию митохондрий и распределение их в различных клетках.
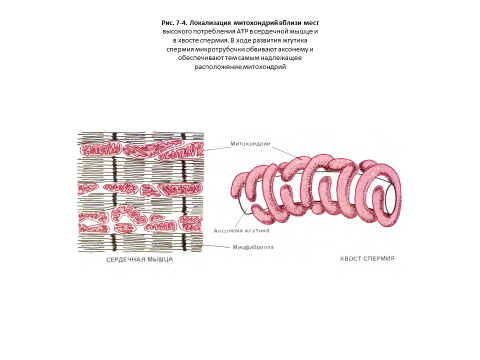
В некоторых клетках митохондрии образуют длинные подвижные филаменты или цепочки, а в других они фиксированы вблизи мест высокого потребления АТР-например, в сердечной мышце они располагаются между миофибриллами, а в сперматозоидах плотно обвивают жгутик (рис. 7-4).
Митохондрии имеют наружную и внутреннюю мембраны, образующие два внутренних компартмента [2]
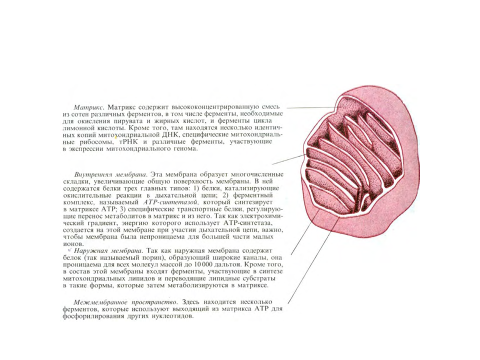
Как видно на рис., каждая митохондрия окружена двумя высокоспециализированными мембранами, играющими ключевую роль в ее активности.
Мембраны образуют два изолированных митохондриальных компартмента: внутренний матрикс и значительно более узкое межмембранное пространство. Если очищенные митохондрии осторожно разрушить и затем разделить на фракции (рис. 7-5), можно определить биохимический состав каждой из двух мембран и заключенных между ними пространств.
Каждая фракция содержит уникальный набор белков.
В состав наружной мембраны входит много копий белка, называемого порином, который образует широкие гидрофильные каналы в липидном бислое. Таким образом, эта мембрана напоминает сито, проницаемое для всех молекул массой 10000 дальтон и меньше, включая небольшие белки. Эти молекулы могут проникать в межмембранное пространство, но большая их часть не способна проходить через непроницаемую внутреннюю мембрану. Это означает, что если химический состав межмембранного пространства эквивалентен составу цитозоля хотя бы в отношении молекул малого размера, то матрикс содержит гораздо более ограниченный набор небольших молекул.
Основная рабочая часть митохондрии - это матрикс и окружающая его внутренняя мембрана. Внутренняя мембрана высокоспецифична, она содержит большое количество «двойного» фосфодипида кардиолипина (разд. 7.5.15), что как полагают, и делает мембрану особенно непроницаемой для ионов. В состав внутренней мембраны входят также разнообразные транспортные белки, обусловливающие ее избирательную проницаемость для тех малых молекул, которые либо метаболизируются многочисленными ферментами, сконцентрированными в матриксе, либо необходимы для их активности. В частности, матрикс содержит ферменты, превращающие пируват и жирные кислоты в ацетил-СоА и затем окисляющие последний в цикле лимонной кислоты. Главные конечные продукты этого окисления - СО2, выходящий из клетки, и NADH, который служит главным источником электронов, переносимых дыхательной цепью - так называется электронтранспортная цепь митохондрий. Ферменты дыхательной цепи встроены во внутреннюю митохондриальную мембрану и необходимы для процесса окислительного фосфорилирования, дающего большую часть АТР в животных клетках.
Энергия, высвобождаемая в процессе переноса электронов по дыхательной цепи, запасается в форме электрохимического протонного градиента на внутренней мембране митохондрий [8]

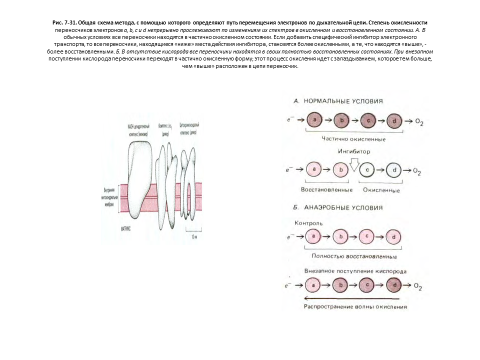

Окислительное фосфорилирование возможно благодаря тесной ассоциации переносчиков электронов с белковыми молекулами. Белки направляют электроны по дыхательной цепи так, что они последовательно переходят от одного ферментного комплекса к другому, не «перескакивая» через промежуточные звенья. Особенно важно то, что перенос электронов сопряжен с аллостерическими изменениями определенных белковых молекул, в результате чего энергетически выгодный поток электронов вызывает перекачивание протонов (Н+) через внутреннюю мембрану из матрикса в межмембранное пространство и далее за пределы митохондрии.
Передвижение протонов приводит к двум важным следствиям:
1) между двумя сторонами внутренней мембраны создается градиент рН - в матриксе рН выше, чем в цитозоле, где значение рН обычно близко к 7,0 (так как малые молекулы свободно проходят через наружную мембрану митохондрии, рН в межмембранном пространстве будет таким же, как в цитозоле);
2) на внутренней мембране создается градиент напряжения (мембранный потенциал), причем внутренняя сторона мембраны заряжается отрицательно, а наружная - положительно.
Градиент рН (?рН) заставляет ионы Н+ переходить обратно в матрикс, а ионы ОН - из матрикса, что усиливает эффект мембранного потенциала (?V), под действием которого любой положительный заряд притягивается в матрикс, а любой отрицательный выталкивается из него. Совместное действие этих двух сил приводит к возникновению электрохимического протонного градиента (рис. 7-19).
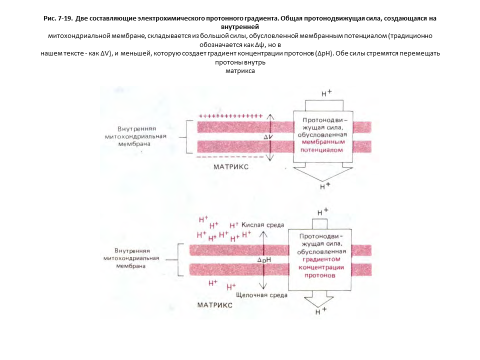
Электрохимический протонный градиент рН (?рН) в 1 единицу рН эквивалентен мембранному потенциалу около 60 мВ, а протонодвижущая сила на внутренней мембране митохондрии составляет около 220 мВ и складывается из мембранного потенциала примерно в 160 мВ и градиента рН, близкого к — 1 единице рН.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
