

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Дыхательная цепь переносит ионы Н+ через внутреннюю митохондриальную мембрану [16]
Если АТР-синтетаза в норме не транспортирует Н+ из матрикса, то дыхательная цепь, находящаяся во внутренней митохондриальной мембране, при нормальных условиях переносит через эту мембрану протоны, создавая таким образом электрохимический протонный градиент, доставляющий энергию для синтеза АТР. При определенных условиях можно экспериментально продемонстрировать способность дыхательной цепи откачивать протоны из матрикса. Можно, например, обеспечить взвесь изолированных митохондрий подходящим субстратом для окисления, а
поток протонов через АТР-синтетазу блокировать. В анаэробных условиях небольшая добавка кислорода к такому препарату увеличит дыхательную активность, которая будет длиться одну-две секунды - пока весь кислород не израсходуется. Во время такой вспышки дыхания с помощью чувствительного рН-электрода можно зарегистрировать внезапное подкисление среды в результате выталкивания ионов Н+ из матрикса митохондрий. Сходный эксперимент можно провести и на взвеси субмитохондриальных частиц. В этом случае при продувании кислорода среда будет подщелачиваться, так как мембрана здесь «вывернута наизнанку» и потому протоны будут накачиваться внутрь частиц.
Перенос электронов осуществляется путем случайных столкновений между донорами и акцепторами электронов, диффундирующими во внутренней митохондриальной мембране [19]
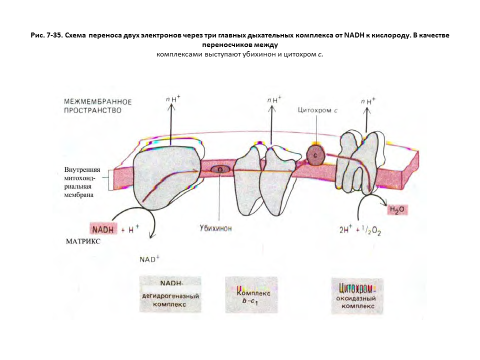
Два компонента, переносящие электроны между тремя главными ферментными комплексами дыхательной цепи, - убихинон и цитохром с
- быстро перемещаются путем диффузии в плоскости мембраны
Дыхательная цепь внутренней митохондриальной мембраны содержит три главных ферментных комплекса, участвующих в переносе электронов с NADH на О2. Если любой из этих комплексов выделить и встроить в мембрану липосомы, то можно продемонстрировать способность его переносить через эту мембрану протоны одновременно с транспортом электронов. В естественной мембране цепь переноса электронов дополняют мобильные переносчики - убихинон и цитохром с, передвигающиеся, подобно челнокам, от одного ферментного комплекса к другому и обратно.
Путь электронов в этой цепи можно представить следующей схемой: NADH > NADH-дегидрогеназный комплекс > убихинон > комплекс b-с1 > цитохром с > цитохромоксидазный комплекс - молекулярный кислород (О2).
Дыхательные ферментные комплексы сопрягают транспорт электронов, сопровождающийся выделением энергии, с откачиванием протонов из матрикса. Создаваемый при этом электрохимический протонный градиент доставляет энергию для синтеза АТР еще одним трансмембранным белковым комплексом-АТР-синтетазой, через которую протоны возвращаются в матрикс. АТР-синтетаза - это обратимый сопрягающий комплекс; в норме он преобразует энергию потока протонов, направленного в матрикс, в энергию фосфатных связей АТР, но при уменьшении электрохимического протонного градиента он способен также использовать энергию гидролиза АТР для перемещения протонов из матрикса наружу.
Отмечу, что хемиосмотические механизмы свойственны как митохондриям и хлоропластам, так и бактериям, что указывает на исключительную важность их для всех клеток.
ФОТОСИНТЕЗ
Всем животным и большинству микроорганизмов необходимо все время получать из окружающей среды большие количества органических веществ. Эти вещества доставляют углеродные остовы для биосинтезов и метаболическую энергию для всех клеточных процессов. Полагают, что первые организмы древней Земли располагали избытком органических соединений, образовавшихся в результате геохимических процессов. Однако большая часть этих соединений была уже использована миллиарды лет назад. С тех времен почти все органические материалы, необходимые для живых клеток, производятся фотосинтезирующими организмами, в том числе разного рода фотосинтезирующими бактериями. Эволюционно наиболее продвинутые из таких бактерий - цианобактерии - обладают минимальными потребностями в питательных веществах. Для превращения атмосферной двуокиси углерода (СО2) в органические соединения они используют солнечную энергию и воду, служащую источником электронов.
Кроме того, при расщеплении воды [при реакции nН2О + nСО2 свет n(СН2О)n + nО2] они выделяют в атмосферу кислород, необходимый для окислительного фосфорилирования в митохондриях. Вероятно, именно эволюция цианобактерии из более примитивных фотосинтезирующих бактерий сделала возможным развитие первых аэробных форм жизни.
У растений, которые появились позже, фотосинтез протекает в специализированных внутриклеточных органеллах - хлоропластах. Однако хлоропласты доставляют энергию для метаболизма только в дневные часы; ночью они прекращают синтез высокоэнергетических метаболитов, и в это время суток растения получают АТР в результате деятельности митохондрий, которые у них очень сходны с митохондриями животных клеток.
Основываясь главным образом на биохимических данных, полагают, что хлоропласты - это потомки цианобактерии, которые захватывались эукариотами путем эндоцитоза и перешли к симбиозу с ними. Так же объясняют и происхождение митохондрий. Согласно этой теории, многочисленные различия между хлоропластами и митохондриями обусловлены отчасти их происхождением от разных бактериальных предков, а отчасти последующей эволюционной дивергенцией. Тем не менее фундаментальные механизмы синтеза АТР в хлоропластах и митохондриях очень сходны, хотя в первых он идет за счет энергии света, а во вторых - за счет энергии, доставляемой дыханием.
Хлоропласты сходны с митохондриями, но имеют один дополнительный компартмент
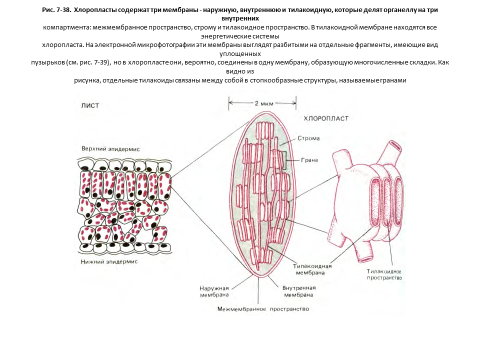
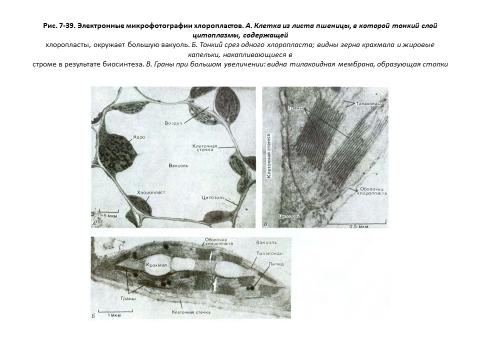
Хлоропласты, так же как и митохондрии, используют для преобразования энергии хемиосмотический механизм, и в основе организациитех и других органелл лежат одни и те же принципы (рис. 7-38 и 7-39). Хлоропласты тоже обладают высокопроницаемой наружной мембраной и гораздо менее проницаемой внутренней, в которую встроены специальные транспортные белки, и эти две мембраны разделены узким межмембранным пространством. Внутренняя мембрана окружает большую центральную область - так называемую строму, представляющую собой аналог митохондриального матрикса и содержащую разнообразные ферменты, рибосомы, РНК и ДНК.
Однако обнаружено и существенное отличие. Внутренняя мембрана хлоропластов не образует крист и не содержит в себе цепи переноса электронов. Фотосинтезирующая поглощающая свет система, электрон-транспортная цепь и АТР-синтетаза находятся в третьей мембране, образующей группу уплощенных дисковидных мешочков - тилакоидов (рис. 7-38). Как полагают, внутренние полости тилакоидов сообщаются между собой, образуя третий, внутренний компартмент хлоропласта, называемый тилакоидным пространством. Это пространство отделено от стромы тилакоидной мембраной.
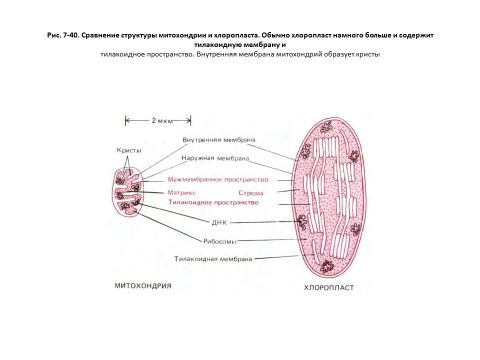
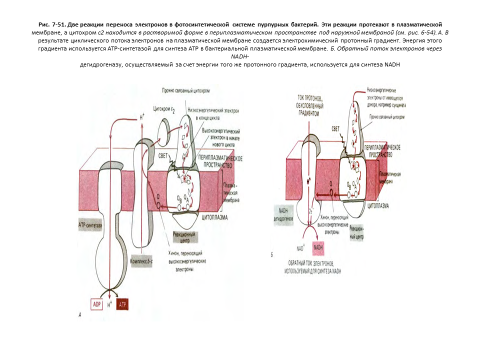
На рис. 7-40 показаны черты сходства и различия в строении митохондрий и хлоропластов. В общем виде хлоропласт можно представить как сильно увеличенную митохондрию, кристы которой образовали в матриксе цепочки связанных между собой субмитохондриальных частиц. В хлоропластах сферическая часть АТР-синтетазы, где образуется АТР, выступает из мембраны тилакоида в область стромы, точно так же как в митохондриях она выступает из внутренней мембраны в сторону матрикса (см. рис. 7-51).
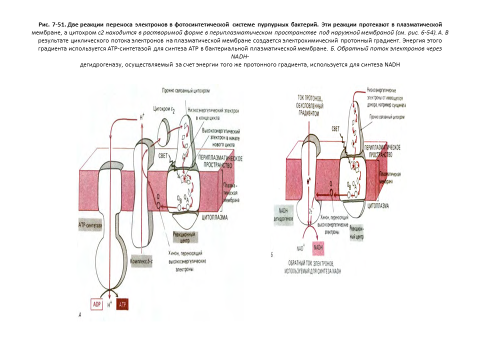
В хлоропластах осуществляются две уникальные реакции: образование АТР и NADPH за счет энергии света и превращение СО2 в углеводы [26]
Разнообразные реакции, протекающие при фотосинтезе, можно разделить на две большие группы.
1. В реакциях фотосинтетического переноса электронов (иногда называемых световыми реакциями) энергия света возбуждает электрон в молекуле хлорофилла, что делает возможным перенос электрона по окислительной цепи в тилакоидной мембране, аналогичный переносу его по дыхательной цепи во внутренней мембране митохондрий. В результате такого транспорта электронов происходит перекачивание протонов через тилакоидную мембрану, и создающаяся протонодвижущая сила доставляет энергию для синтеза АТР в строме.
В митохондриях, электроны, образуемые в окислительной цепи, восстанавливают NADP+ до NADPH. Источником электронов, участвующих в этом процессе, служит окисление воды, при котором выделяется О2.
2. В реакциях фиксации углерода (называемых иногда темновыми реакциями) происходит превращение СО2 в углеводы, причем в качестве источника энергии и восстанавливающего агента используются соответственно АТР и NADPH, синтезированные в реакциях фотосинтетического переноса электронов. В результате этих реакций, начинающихся в строме хлоропласта и продолжающихся в цитозоле, в листьях образуется сахароза, откуда она доставляется к другим частям растения, где служит источником энергии для роста и используется при синтезе органических молекул.
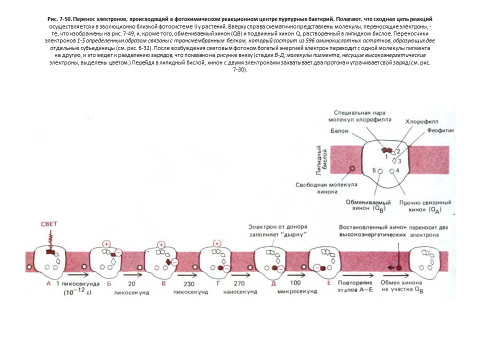
Фотосинтез определяется фотохимией молекулы хлорофилла [31]
Рассмотрев реакции связывания углерода, вернемся теперь к вопросу о том, как в процессе фотосинтетического переноса электронов, протекающем в хлоропласте, образуются АТР и NADH, необходимые для синтеза углевода из СО2 и Н2О.

Необходимая энергия извлекается из солнечного света, поглощаемого молекулами хлорофилла (рис. 7-46). Процесс преобразования энергии начинается с возбуждения молекулы хлорофилла квантом света (фотоном), сопровождающегося переходом электрона на более высокий энергетический уровень. Такая возбужденная молекула нестабильна и стремится вернуться к исходному состоянию одним из трех способов:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
