

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Фотосинтез осуществляется за счет перехода возбужденных электронов с Т на Т*, при котором изменяется их спин. Появление же неспаренных электронов приводит к образованию свободных радикалов. Возникновение свободных радикалов при фотосинтезе доказано методом ЭПР.
Полный цикл фотосинтетических превращений начинается с фотовозбуждения хлорофилла а в ФС II (Р680). Возбужденный электрон переносится с Р680 на акцептор, тесно контактирующий с хлорофиллом в тилакоидной мембране. По-видимому, акцептором служит цитохром (Q). На переход затрачивается энергия возбуждения электрона, так как он преодолевает восстановительный потенциал в 1,24 В (идет «в гору»). Приняв электрон, акцептор восстанавливается, а в молекуле хлорофилла, отдавшей его, возникает дырка. Она заполняется электроном воды, которая расщепляется в процессе фотосинтеза с образованием кислорода, протонов и электронов. По-видимому, расщепление воды (точнее, ее окисление до О2) производит не сам возбужденный хлорофилл, а некое промежуточное окисленное соединение, образующееся при возбуждении Р680. Окисление воды, приводящее к выделению кислорода, катализируется ферментной системой, содержащей трехвалентный ион марганца.
Присоединив электрон, хлорофилл (Р680) восстанавливается, и к нему возвращается способность возбуждаться новыми квантами света. Кислород выделяется в атмосферу. Протоны закисляют содержимое тилакоидов, поддерживая там более высокую концентрацию Н+ по сравнению с цитозолем. Такова судьба всех продуктов расщепления воды при фотосинтезе.
Теперь обратимся к электронам, покинувшим молекулы хлорофилла (Р680) и восстановившим вещество Q. Проследим их миграцию по цепи переноса ФС II. С вещества Q они последовательно переходят от одного компонента транспортной цепи к другому в направлении более высокого положительного восстановительного потенциала («с горы») –к пластохинону и далее к пластоцианину. Свободная энергия, выделяющаяся при этом ступенчато (дискретно), запасается тилакоидной мембраной и обеспечивает синтез АТФ из АДФ и ортофосфата. Так как скорость переноса электронов по транспортной цепи ФС II не зависит от температуры, то возникло предположение о туннельном механизме миграции электронов между окислительно-восстановительными компонентами этого каскада.
Последним из них в ФС II является Р700, т. е. хлорофилл а, принадлежащий другой фотосистеме (ФС I) в данной квантосоме. Однако Р700 приобретает способность принять электроны из ФС II только после того, как сам поглотит свет, возбудится и потеряет свой электрон, который поступит в транспортную цепь ФС I. При потере электрона в молекуле Р700 возникает дырка, которую и занимает электрон, пришедший сюда из ФС II (с пластохинона). Следовательно, возбужденный хлорофилл, входящий в состав ФС II, восстанавливает пластохинон (до гидрохинона), а возбужденный хлорофилл, принадлежащий ФС I, окисляет гидрохинон до пластохинона, возвращая ему способность принимать новые электроны от Р680. Такое сопряжение в работе двух фотосистем осуществляется в пределах одной квантосомы. В ней между молекулами хлорофилла, принадлежащими разным фотосистемам, иными словами, между Р680 и Р700, расположено примерно 5 молекул пластохинона. Пигмент Р700, приняв электроны от компонентов ФС II, восстанавливается, и к нему возвращается способность возбуждаться светом.
В ФС I пока не идентифицировано вещество с наибольшим (по модулю)
отрицательным восстановительным потенциалом, на которое переходят возбужденные электроны с Р700. Обозначим его буквой X. Приняв электрон от возбужденного хлорофилла, X становится сильным восстановителем (Х-), с которого начинается второй каскад переноса электрона в сторону более высокого положительного восстановительного потенциала.
Последним компонентом цепи транспорта электронов в ФС I является НАДФ+, восстанавливаемый до НАДФН. Донором водорода служит вода, расщепившаяся под действием света при участии хлорофилла (Р680) еще на первой стадии фотосинтеза.
Конечным акцептором электронов, прошедших последовательно ФС II и ФС I, является СО2. Перенос на него электронов с редокс-пары НАДФ+/НАДФН
(восстановительный потенциал равен -0,324 В) осуществляется сложной ферментной системой, входящей в так называемый цикл Кальвина – последовательность химических реакций, протекающих в 13 стадий. Энергия, выделяемая при транспорте электронов с названной редокс-пары на СО2, недостаточна для его восстановления до углеводов.
Дополнительная энергия, необходимая для синтеза углеводов, поступает в цикл Кальвина в форме АТФ. Так, образование одной молекулы гексозы из 6 молекул СО2 обеспечивается не только переносом электронов с НАДФН на СО2, но еще и 18 молекулами АТФ. Этот макроэрг образуется также в реакциях фотосинтеза посредством преобразования солнечной энергии, поглощенной хлорофиллом. Синтезу АТФ служит транспорт в квантосоме (в ФС II и ФС I) до этапа восстановления НАДФ+.
За счет поглощения двух квантов света двумя молекулами хлорофилла (Р680 и Р700), принадлежащими одной квантосоме, происходит перенос одного электрона на СО2. Кроме того, энергия этих двух фотонов обеспечивает перенос одного электрона с воды на Р680.
Рассматривая процессы электронного транспорта в ФС II и ФС I в совокупности, можно заключить, что поглощение двух квантов света двумя молекулами хлорофилла приводит к переносу одного электрона с Н2О на СО2, из которого синтезируются углеводы. В ходе двухтактного (в ФС II и ФС I) фотоэлектронного преобразования возникают молекулярный кислород, НАДФН (восстановленный НАДФ), а также АТФ. Это единственная, хотя и сложная (многоступенчатая) световая реакция в процессе фотосинтеза.
Дальнейшие аутотрофные биосинтетические реакции не нуждаются в свете. Они происходят в зеленых растениях за счет энергии, заключенной в электронах, принадлежащих НАДФН и АТФ. Лучше других реакций фотосинтеза изучен процесс образования молекулы глюкозы из 6 молекул СО2 и 6 молекул Н2О (реакция фиксации углекислого газа). 12 пар возбужденных электронов, необходимых для осуществления этой реакции, поступают от НАДФН. Дополнительным поставщиком свободной энергии служат 18 молекул АТФ. Вклад НАДФН и АТФ в фотосинтез одного моля глюкозы составляет соответственно 660 и
252 ккал.
Число митохондрий и хлоропластов в клетке поддерживается путем их деления [46]
Митохондрии и хлоропласты никогда не возникают de novo, они всегда образуются путем деления уже существующих органелл. Как показывают наблюдения над живыми клетками, митохондрии не только делятся, но могут и сливаться друг с другом. Однако в среднем каждая органелла должна удвоить свою массу и затем разделиться пополам один раз за одну клеточную генерацию. Электронные микрофотографии дают основание полагать, что деление митохондрий начинается с образования кольцевой бороздки на внутренней мембране, подобно тому как это происходит при делении многих бактериальных клеток (рис. 7-65 ; таким образом, деление митохондрий - это, по-видимому, контролируемый процесс, а не случайное расщепление надвое.
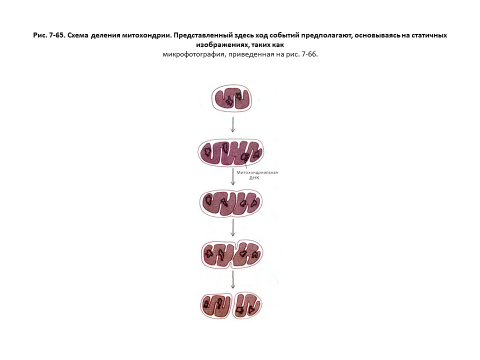
В большинстве клеток энергопреобразующие органеллы делятся на протяжении всей интерфазы; таким образом, каждая из них делится независимо от остальных и от всей клетки. Точно так же репликация ДНК в органеллах происходит не только в период синтеза ядерной ДНК (S - фаза), но и в другие фазы клеточного цикла. Хотя, по-видимому, индивидуальные молекулы ДНК реплицируются случайным образом (так что в данном клеточном цикле одни могут удвоиться несколько раз, а другие ни разу), общее число их за каждый клеточный цикл удваивается, поддерживая постоянство количества этой ДНК в клетке. Число энергетических органелл может регулироваться в зависимости от потребности клетки в энергии; например, значительное увеличение (в 5-10 раз) количества митохондрий наблюдается при многократном сокращении скелетной мышцы в течение длительного периода.
Более того, в ряде случаев деление органелл регулируется клеткой: так, хлоропласты некоторых водорослей, содержащих только одну или несколько таких органелл, делятся непосредственно перед цитокинезом, причем в той же плоскости, в которой будет происходить очередное деление клетки. Но действующие при этом регуляторные механизмы на молекулярном уровне не изучены.
В большинстве случаев геномы хлоропластов и митохондрий представлены кольцевыми молекулами ДНК [47]
Молекулы ДНК органелл относительно просты, невелики и (за исключением митохондриальных геномов некоторых водорослей и простейших) замкнуты в кольцо. Размеры генома хлоропластов у всех исследованных организмов сходны, тогда как митохондриальные геномы_отличаются._
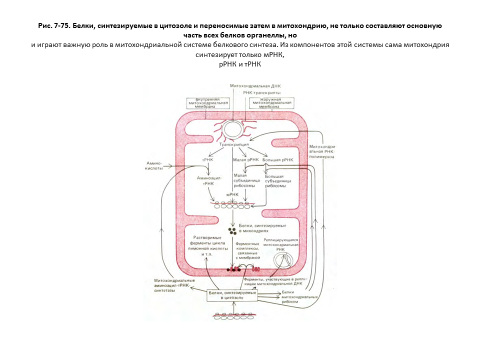
Несмотря на небольшое число белков, кодируемых генами митохондрий и хлоропластов, эти органеллы осуществляют репликацию и транскрипцию своей ДНК и белковый синтез. Эти процессы протекают в матриксе митохондрий и строме хлоропластов. Хотя белки, участвующие
во всех этих процессах, специфичны для органелл, большая часть их кодируется ядерным геномом (разд. 7.5.17). Это тем более удивительно в связи с тем, что весь аппарат белкового синтеза в органеллах сходен с бактериальным, а не с эукариотическим. У хлоропластов это сходство особенно велико:
1. Рибосомы хлоропластов очень напоминают рибосомы Е. coli как по своей чувствительности к различным антибиотикам (хлорамфениколу, стрептомицину, эритромицину, тетрациклину и др.), так и по структуре. При этом не только поразительно сходны нуклеотидные последовательности рибосомных РНК хлоропластов и Е. coli, но рибосомы хлоропластов способны использовать тРНК бактерий при синтезе белка. Во всех этих отношениях рибосомы хлоропластов отличаются от рибосом, находящихся в цитозоле растительных клеток.
2. Синтез белка в хлоропластах начинается с N-формилметионина, как и у бактерий, а не с метионина, как в цитозоле эукариотических клеток.
3. ДНК хлоропластов в отличие от ядерной ДНК может транскрибироваться с помощью РНК-полимеразы из Е. coli с образованием хлоропластных мРНК, которые эффективно транслируются белок-синтезирующей системой Е. coli.
Хотя у митохондрий генетические системы гораздо менее сходны с аналогичными системами современных бактерий, чем у хлоропластов,
ИТАК
Рост и деление митохондрий и хлоропластов контролируются двумя отдельными генетическими системами: геномом самой органеллы
и ядерным геномом. Большая часть белков этих органелл закодирована в ядерной ДНК, синтезируется в цитозоле и затем переходит в органеллу. Однако некоторые белки митохондрий и хлоропластов и все их РНК
кодируются в ДНК самих органелл и в них же синтезируются. Геном митохондрий человека содержит около 16500 пар нуклеотидов и кодирует
рибосомные РНК, 22 транспортные РНК и 13 различных полипептидных цепей. Геном хлоропластов примерно в 10 раз больше генома митохондрий человека и содержит около 120 генов. Однако преобладающая роль в биогенезе органелл обоих типов принадлежит ядру: это подтверждается тем, что даже у таких мутантов, у которых отсутствует функционирующий геном органелл, частично функционирующие органеллы образуются в нормальном количестве.
Рибосомы хлоропластов очень сходны с бактериальными рибосомами, тогда как рибосомы митохондрий несколько больше отличаютсяот последних; поэтому проследить происхождение митохондрий сложнее. Однако сходство между белками дает основание предполагать, что те. и другие органеллы произошли от бактерий, вступивших в устойчивый симбиоз (в качестве эндосимбионтов) с какими-то примитивными эукариотическими клетками; как полагают, митохондриям дали начало пурпурные бактерии, а хлоропластам (позднее) - цианобактерии или близкие к ним организмы. Хотя многие гены этих древних бактерий все еще используются для синтеза белков органеллы, большая их часть по неясным причинам включилась в ядерный геном, где они кодируют ферменты, которые сходны с бактериальными и синтезируются на рибосомах в цитозоле, а затем переходят в органеллу.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
