

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
(Рис. 6. Динамика числа КОЕ при культивировании при комнатной температуре в начальный период таяния образцов, а также через 5, 7 и 10 суток после оттаивания (подсчет числа колоний осуществляли на 5-ые сутки культивирования))
Численность культивируемых гетеротрофных бактерий, высеянных из образцов в начальный период таяния, через 7 и 14 суток после оттаивания различается по трем основным группам образцов (см. рис. 7 ):
1) каменная мостовая, в которой первичные продуценты ОВ представлены гиполитными сообществами зеленых водорослей и цианобактерий, преимущественно в виде биопленок (обр. 10, 13),
2) моховые подстилки и горизонты с примесями оторфованного материала (обр. 1d, 3, 14, 5d) и
3) подстилающие минеральные горизонты B (обр. 15, 3d, 4d).
Низкая численность гетеротрофных бактерий ожидаемо обнаружена в минеральных песчаных горизонтах B, для которых характерно небольшое содержание органического углерода. Из этой группы наибольшая численность наблюдалась в горизонте Bfungi (обр. 3d) c мицелиальными грибами, а, возможно, и актиномицетами. Повышенная численность бактерий и углерода характерна для моховых подстилок и оторфованного материала. Наиболее интересным является образец каменной мостовой, в которой в поле отсутствовали макропризнаки биоты, но оказалась высокая численность бактерий при высеве через 7 и 10 суток после оттаивания образца при низком содержании органического углерода. По данным СЭМ органическое вещество здесь представлено цианобактериальными биопленками и аморфными органоминеральными пленками (1-8 микрон) сложного генезиса на поверхности частиц кварца и полевых шпатов.
Наиболее распространенной группой растений, представленной в изучаемых оазисах Восточной Антарктики являются высшие растения бриофиты - мхи. ОВ мохового генезиса - это более сложные органические соединения, нуждающиеся» в повышенной ферментативной активности для их деструкции. Другие первичные продуценты ОВ в антарктических оазисах, хлорофиты и цианофиты (зеленые водоросли и цианобактерии) организованы преимущественно в виде биопленок с привлечением защитных механизмов в виде внеклеточного полимерного матрикса. По-видимому, их деструкция связана с иным уровнем активности, а возможно и составом ферментов.
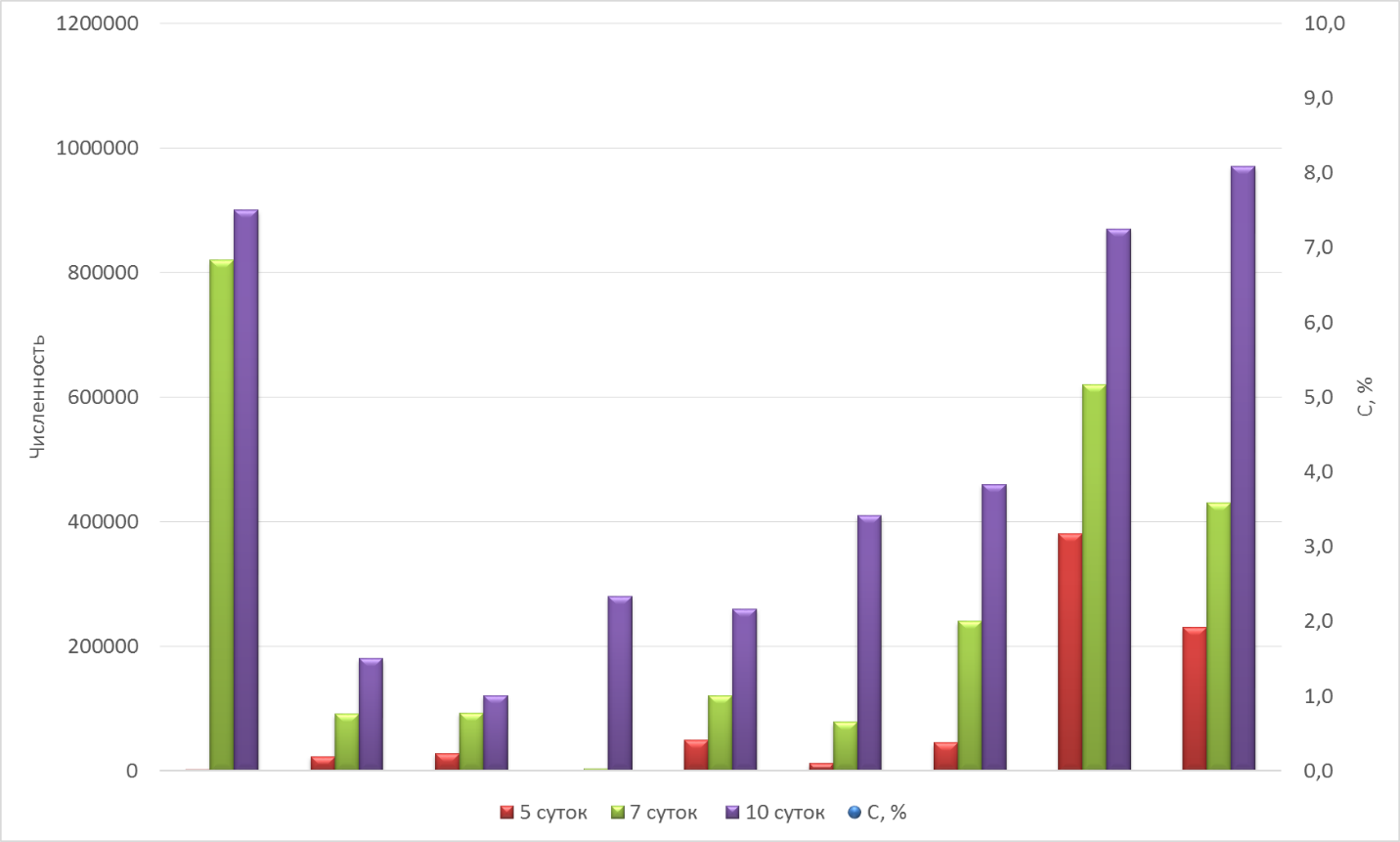
(Рис. 7. Сравнительные показатели динамики числа КОЕ при высеве из образцов различных горизонтов формирующихся почв в разные сроки после их оттаивания.)
Как показало изучение каталазной и дегидрогеназной активности в исследованных образцах их активность была низкой. В некоторых образцах она не обнаруживалась в начальный период оттаивания (№ 13 и 15).
В целом, изучение динамики каталазной и дегидрогеназной активности ферментов после первичного оттаивания образцов, и далее через 7 и 14 суток после оттаивания выявило следующее:
Динамика активации каталазы в процессе хранения образцов после их оттаивания выявила существенный разброс по показателям в сразу же после оттаивания и в последующие 7 и 14 суток после оттаивания - в некоторых образцах она возрастала к 7-м суткам 5Д, 3Д и 14), а в дальнейшем падала, а в образце наоборот возрастала на 14 сутки после оттаивания. Т. е. в разный период оттаивания показатели менялись, и это изменение не коррелировало с возрастающим числом КОЕ.
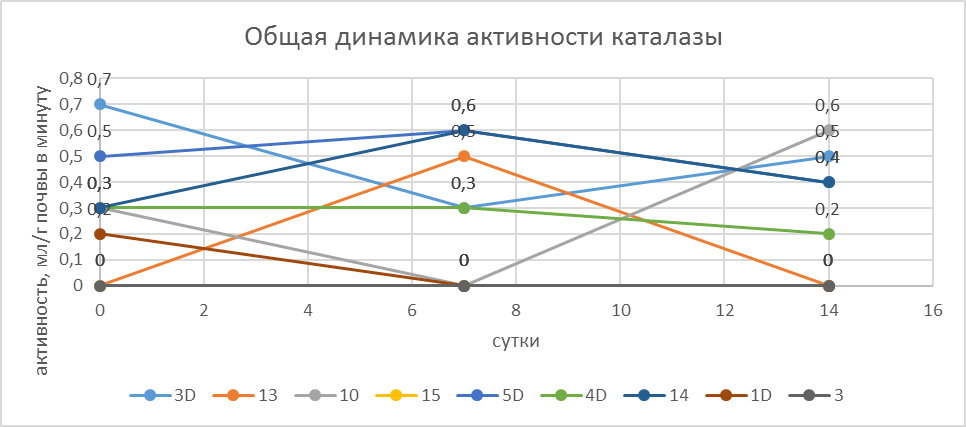
(Рис. 8. Динамика активности каталазы в разные сроки после оттаивания.)
Если автивность каталазы проявлялась прктически во всех образцах, активность дегидрогеназы была также низкой и в большинстве образцов не выявлялась, как сразу после оттаивания образцов, так и через 10 и 14 суток хранения при комнатной температуре при одновременно увеличении числа КОЕ.
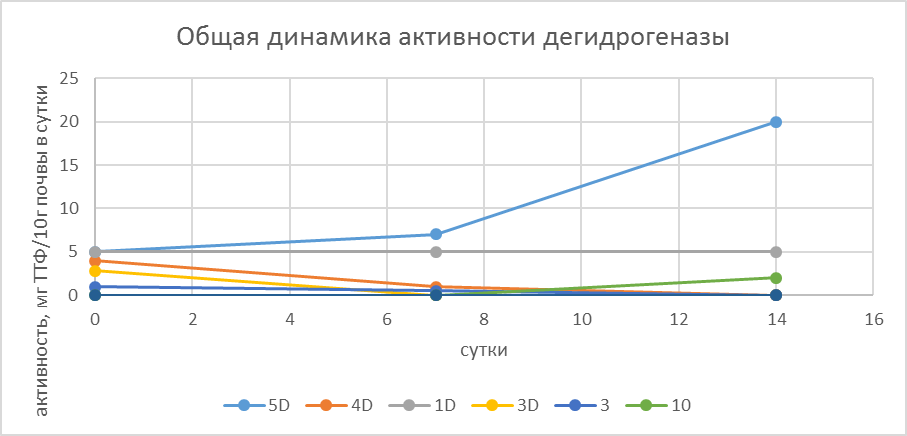
(Рис. 9 Динамика активности дегидрогеназы в разные сроки после оттаивания.)
Несмотря на низкие показатели активности каталазы и дегидрогеназы в исследованных горизонтах примитивных почв, можно сделать некоторые выводы, которые даже при низких значениях могут охарактеризовать изученные образцы.
Наиболее высокая каталазная активность наблюдалась в горизонте Bfungi (обр. 3d), в начальный период оттаивания – до 0.7 мл О2 / г в мин, некоторых моховых подстилках и оторфованном материале до 0.5 мл О2 / г в мин, а также в некоторых горизонтах каменной мостовой до 0.3 мл О2 / г в мин.
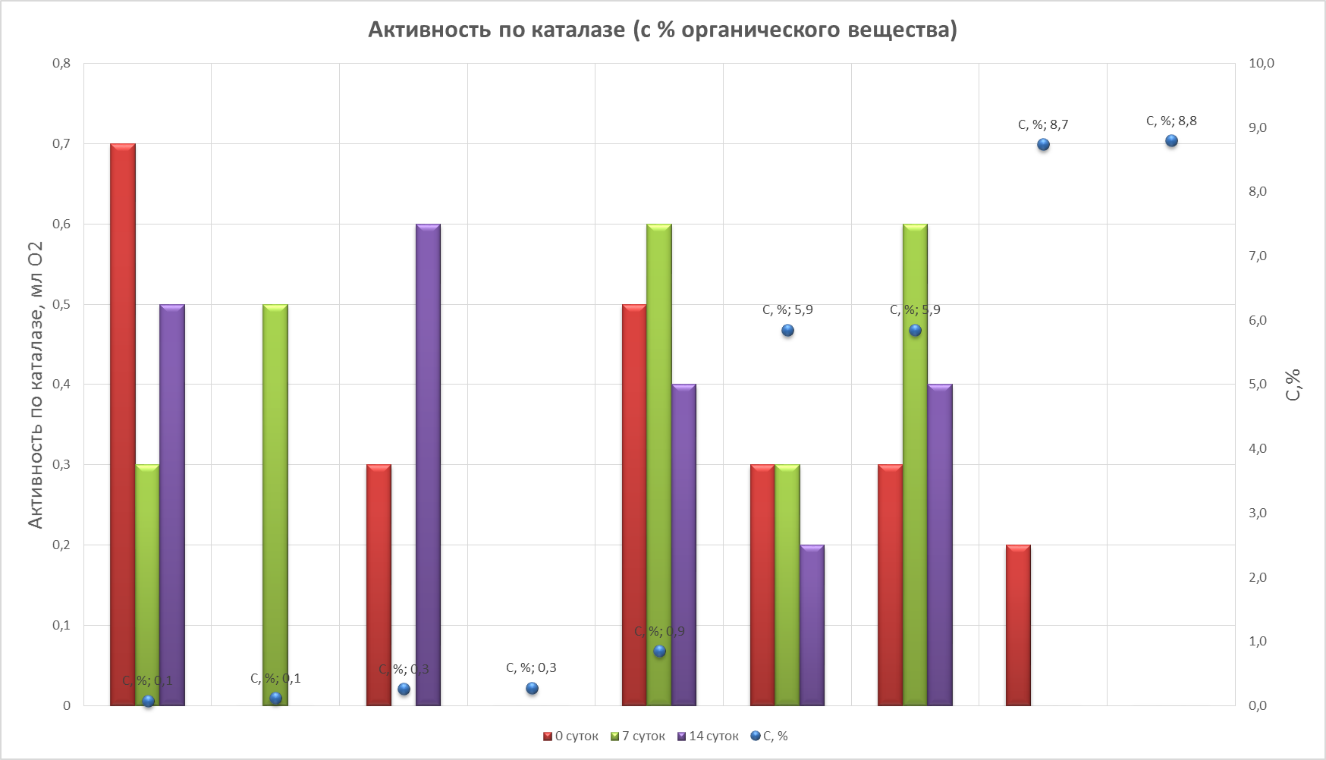
(Рис.10. Активность по каталазе.)
Наибольшую дегидрогеназную активность имеют некоторые горизонты, где есть свежие или мертвые ткани мха. В некоторых моховых подстилках и одном горизонте каменной мостовой активность по ходу эксперимента нарастала или оставалась одинаковой. В горизонтах B, наоборот, снижалась.
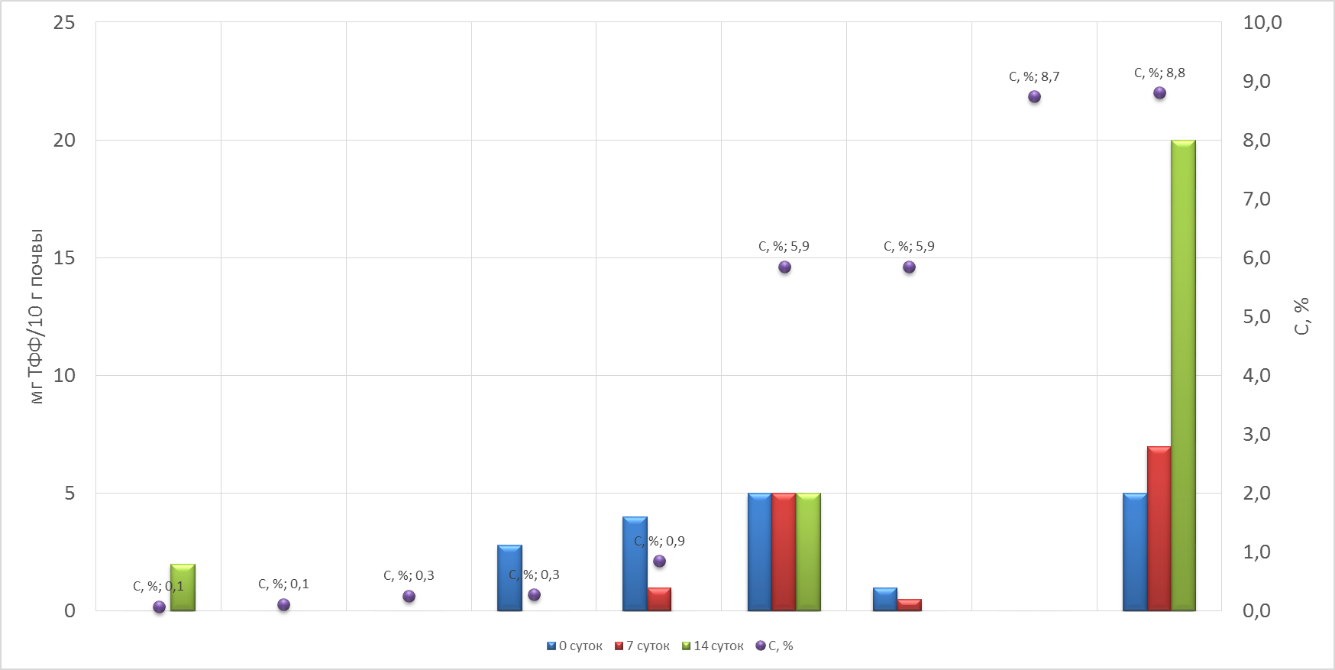
(Рис. 11. Активность по дегидрогеназе.)
2.2.4.Сравнительный морфологический анализ культивируемых бактерий.
Морфотипы выделенных бактериальных культур определяли по цвету и форме колоний, из которых в последующем готовили препараты для световой микроскопии и изучали их морфологию. Выделенные в чистую культуру штаммы относили предварительно к грамотрицательным или грамположительным формам по тесту с КОН.
Среди выделяемых из поверхностных горизонтов бактерий - образцов каменной мостовой (№ 5Д, 10), песчаного мелкозема в наскальной ванне под сообществом эпилитных лишайников (4Д), моховой подстилки с минеральными примесями (№ 3) большинство выделенных культур относились к окрашенным формам - оранжевым, желтым, ярко - розовым, что по-видимому, позволяет им противостоять сильному ультрафиолетовому излучению, которое характерно для оазисов Антарктиды. В нижележащих горизонтах увеличивалось число неокрашенных форм. Обращает на себя внимание тот факт, что выделяемые на чашках после первоначального высева культуры по культурально-морфологическим признакам могут быть отнесены к небольшому числу таксонов, в отличие от культур, выделяемых из арктических почв, а также антарктических мерзлых осадков.
Среди культивируемых культур споробразующие бактерии были выделены только из образца торфянистого горизонта (№ 13). Редкое выделение спорообразующих бактерий вообще характерно для местообитаний с отрицательными температурами, в частности, для нижележащих мерзлых осадков (Vorobyova et al., 1997, 2001). Обращает на себя внимание, также, присутствие в бактериальном сообществе изученных антарктических почв стрептомицетов, которые выделяются крайне редко из нижележащих горизонтов мерзлых осадков, но в тоже время обнаруживаются в образцах антарктических льдов.
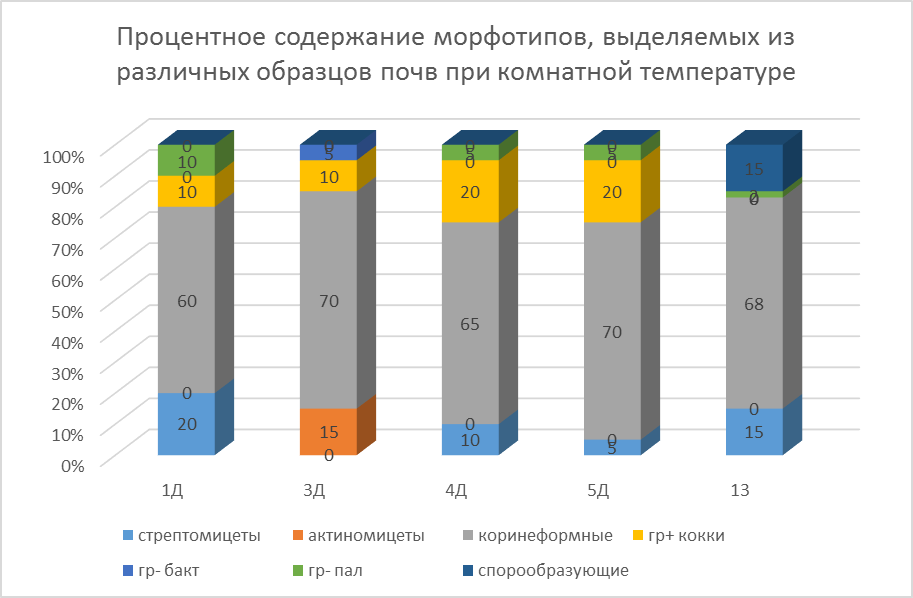
(Рис.12. Процентное содержание морфотипов, выделяемых из различных образцов почв при комнатной температуре.)
По морфологии выделенные нами культуры были разделены на 4 типа: коринеформные, грамположительные кокки, стрептомицеты и палочки с грамотрицательным типом клеточной стенки. На рис. 12. видно, что большинство культур относится к коккам и коринеформным бактериям, а грамотрицательные палочки составляют меньшинство.
Чередующиеся циклы замораживания - оттаивания выявили, что в большинстве случаев изменений в морфотипах выделяемых культур не наблюдалось, за исключением образца почвы, формирующейся под моховым сообществом в днище влажной долины (№1Д) и образца песчаного мелкозема в наскальной ванне под сообществом эпилитных лишайников. (№4Д), где выделялись спорообразующие бактерии.
Функциональное разнообразие бактериальных сообществ.
Разнообразие потребляемых субстратов мало изменяется по профилю глеезема и органоминеральной почвы с поверхностным водорослевым горизонтом, в то время как оно снижается по профилю почвы под каменной мостовой.
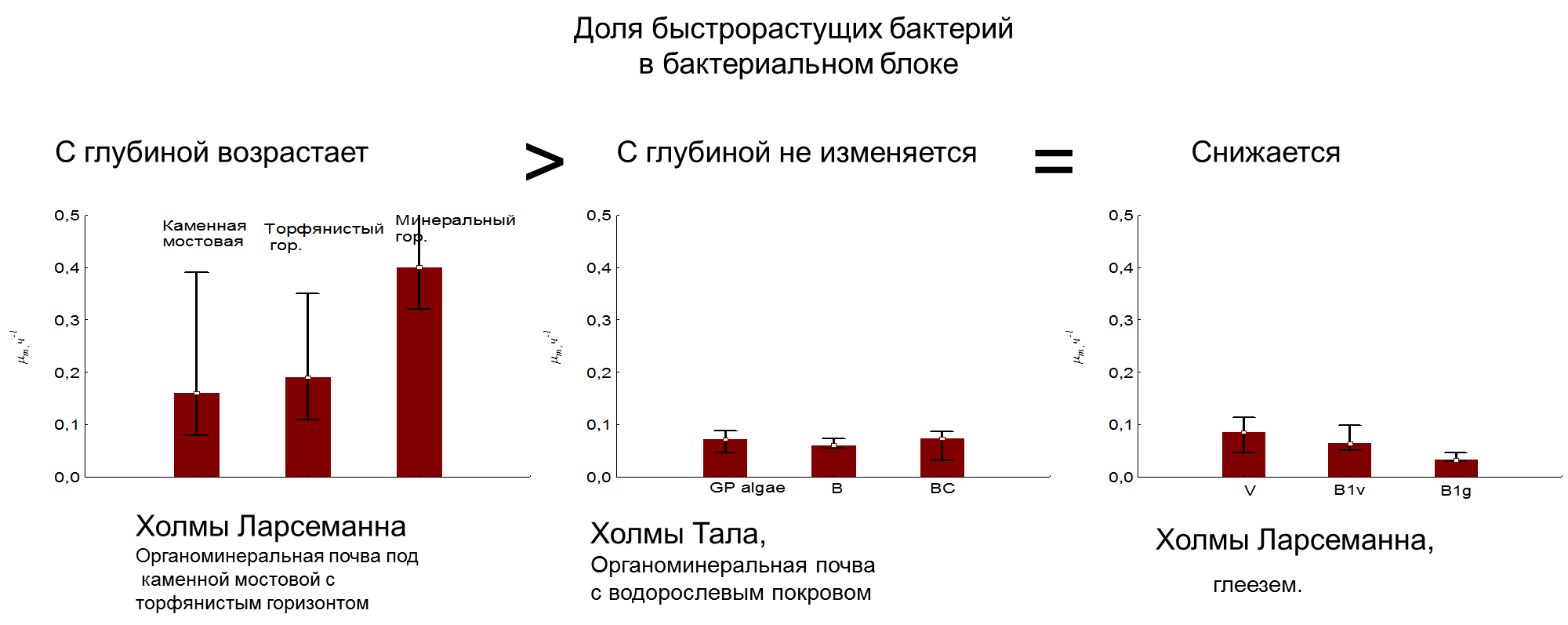
(Рис 13. Доля быстрорастущих представителей в бактериальном блоке)
Однако, доля быстро растущих бактерий в профиле этой почвы наоборот увеличивается (рис 13). Другие типы почв обнаружили иную тенденцию: либо с глубиной не изменяется данный показатель (почвы Холмов Тала) либо снижается – в глееземе оазиса холмов Ларсеманна. Это объясняется тем, что доля быстрорастущих ассоциаций отражает долю быстрорастущих бактерий (r-стратегов) в сообществе почв, а доля быстро растущих бактерий в профиле этой почвы наоборот увеличивается.
Таблица 5. Физиологическое состояние образцов.
Субстрат | Горизонт | % потребляемых субстратов, | % ячеек в которых есть рост | Субстраты |
Холмы Талла, органоминеральная с водорослевым покровом | GP algae 0-1см | 42 | 39 | Казеин, твин, НК. Ксилан, крахмал |
B 4-7см | 33 | 33 | Казеин, твин, НК., крахмал | |
BC 20-25см | 25 | 25 | Казеин, твин, НК | |
Холмы Ларсеманна, глеезем | V (0-4 см) | 67 | 44 | Казеин, твин, НК кератин, ксилан крахмал декстан пектин |
B1v (4-17см) | 42 | 31 | Казеин, твин, НК хитин, ксилан | |
B1g (4-17см) | 67 | 58 | Казеин, твин, НК кератин, ксилан крахмал пектин, хитин | |
Холмы Ларсеманна, органоминеральная с торфянистым горизонтом под каменной мостовой | Каменная мостовая (0-2см) | 87 | 65 | КМЦ крахмал казеин, крахмал, хитин ксилан декстран |
Торфянистый горизонт (2-4см) | 75 | 53 | Казеин, твин. крахмал, хитин ксилан декстран | |
Минеральный горизонт (4-10 см) | 37 | 30 | Крахмал, казеин хитин |
Доля метаболически готовых к росту на полимерах ассоциаций (воспринимаемые нами как целое), отражает действительную метаболическую готовность к потреблению полимеров in situ (Таблица 5.).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
