

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Профиль II - образцы почвы с моховой подстилкой, развивающейся под щебнистой броней каменной мостовой, которые появляются, начиная со значений близких к нижнему пределу полной влагоемкости песчаных почв (~18%), и исчезают, когда влажность поверхностного горизонта падает ниже 9-10%, то есть приближается к наименьшей влагоемкости песчаного субстрата.
Профиль III - образцы почвы, где из визуальных признаков жизни имеется лишь оливковый органоминеральный горизонт, образовавшийся за счет развития колоний водорослей в приповерхностном песчаном материале. Его влажность составляет 4-8%, то есть остается в пределах нижней части диапазона доступной для организмов влаги в песчаных почвах. Это зона слабого влияния снежника
Профиль IV - образец верхнего слоя безгумусового почвоподобного тела, развивающегося на высоких бортах долин и имеющего влажность всего лишь 0,5-2%, то есть близкую к гигроскопической для крупнопесчаных почв.
Определение численности бактерий в образцах тундровой почвы и мерзлых отложений проводили чашечным методом и методом прямого счета. Численность бактерий в образцах определяли прямым микроскопическим люминисцентным методом с использованием красителя акридина оранжевого ("Методы почвенной микробиологии и биохимии", 1980).
Для определения численности жизнеспособных бактерий чашечным методом для посева готовили водную суспензию с разведением 1:10 и высевали на агаризованную триптиказо-соевую среду (ТСА)
Рассев каждого образца делали в трехкратной повторности. Подсчет числа колониеобразующих единиц (КОЕ) производили после инкубации от трех до 15 суток при комнатной температуре.
Активизацию роста культивируемых бактерий изучали по динамике численности бактерий в процессе размораживания образцов почв по числу КОЕ, выявляемому при культивировании на ТСА при +200 С, сразу же после оттаивания образцов, а также через 5, 7. и 14 суток. Контролем служила численность бактерий, определенная по числу КОЕ на ТСА и температуре культивирования при выделении из нативных образцов почв сразу же после оттаивания.
Для изучения влияния процессов замораживания - оттаивания на жизнеспособность бактерий in situ образцы почвы, хранившиеся в морозильной камере при –180С, подвергались оттаиванию при комнатной температуре (+23 0С) в течение 1 часа и последующим чередующимся циклам замораживания (в морозильной камере при –180 С) в течение 3-х суток) и оттаивания при тех же условиях, что и нативные образцы. После 10 и 30 чередующихся циклов замораживания - оттаивания определяли жизнеспособность бактерий по числу КОЕ на триптиказо-соевой среде через 5 суток культивирования при комнатной температуре. Контролем служило число КОЕ, полученное после первоначального оттаивания нативных образцов.
О качественном составе бактерий, выявляемых на ТСА сразу после оттаивания образцов и последующих циклов замораживания – оттаивания судили по морфологическим особенностям клеток, наблюдаемым в фазово-контрастном микроскопе. Принадлежность выделенных культур к бактериям с грамотрицательным и грамположительным типами клеточных стенок предварительно судили по тесту с КОН.
Определяли динамику каталазной и дегдрогеназной активности исследуемых образцов почв в начальный период таяния и через 7 и 14 суток после их оттаивания общепринятыми методами – газометрическим для каталазы и калориметрическим методом для дегидрогеназы («Методы почвенной микробиологии и биохимии», МГУ, 1980.
Функциональное разнообразие бактериальных комплексов изучали в профиле эндолитной почвы с торфянистым горизонтом и профиле глеезема, формирующегося в сапропеле пресноводного озера (Холмы Ларсеманна. станция Прогресс); а также органоминеральной почве с водорослевым покровом, отобранной в районе оазиса Холмы тала (станция Молодежная).
Также определяли их максимальную удельную скорость роста на питательных средах, содержащих различные органические вещества. Использовали метод, основанный на определении продолжительности лаг-фазы роста микроорганизмов, выделяемых из природных субстратов на питательные среды, которая может зависеть не только от условий культивирования, но и от физиологического состояния микроорганизмов в природных субстратах перед их выделением. Предполагается, что чем короче лаг фаза роста микроорганизма в питательной среде, тем в более активном состоянии находился организм в природных условиях. Способность к активному росту микроорганизмов определялась по кривой начального роста на питательных средах.
В основе метода лежит синтетическая хемостатная модель, упрощенная для описания лаг-фазы и фазы нелимитированного роста периодической культуры микроорганизмов и рассчитывается по уравнению
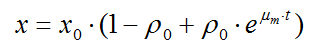
Применяемый метод позволяет также определить функциональное (трофическое) разнообразие микробных сообществ (число сред, на которых регистрируется рост) и кинетический параметр роста - максимальную удельную скорость роста, которая отражает экологическую стратегию роста бактерий.
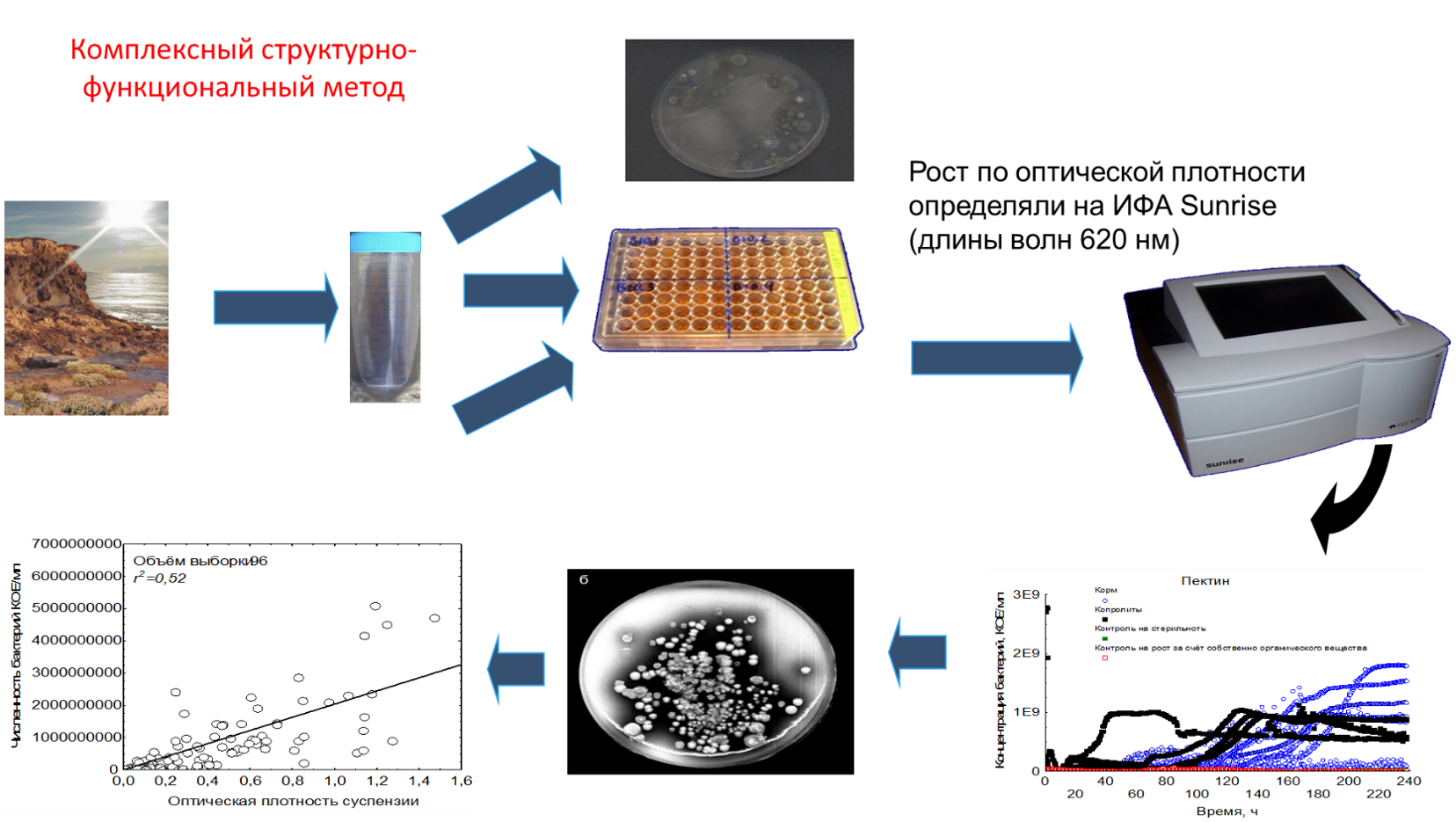
(Рис 4. Инфографика комплексного структурно-функционального метода.)
Методика определения следующая:
Водная суспензия исследуемых объектов вносилась в ячейки культуральной планшеты, с внесёнными в них биополимерами животного, растительного и микробного происхождения. Кривые периодического роста возникших на полимерах бактериальных ассоциаций детектировали по оптической плотности сред. На основании калибровочной прямой производился пересчёт оптической плотности в концентрацию клеток в среде. Кинетика роста ассоциаций на полимерах описывалась математической моделью периодической культуры.
В данной работе теоретически интерпретировался только участок подготовки микроорганизмов к росту (лаг-фаза) и фаза экспоненциального роста на основе новой комплексной модели периодической культуры , которая для этих стадий роста очень близка к синтетической хемостатной модели . Эта модель может быть записана в форме системы дифференциальных уравнений или в виде одного интегрального уравнения. В модели экспериментальные данные это X – концентрация клеток в среде, X нулевое - исходная концентрация в суспензии культивируемых бактерий и T (время), а параметры мю м и ро. Мю - это удельная скорость роста микроорганизмов, то есть скорость роста единицы обилия микроорганизмов (биомассы, численности). Мю м - максимальная удельная скорость роста. Ро - функция физиологического состояние растущей культуры, отражающее отличие в лаг-фазе удельной скорости роста мю от мю м. Параметр ро нулевое характеризует физиологическое состояние в момент интродукции (то есть в природе). Метаболическая готовность к росту (гамма) – это фактически готовность микроорганизма к росту на конкретной среде. Мы полагаем, что метаболическая готовность к росту определяется физиологическим состоянием. Однако поскольку значения Ро нулевого колеблются на несколько порядков то необходимо логарифмировать эту величину и прибавить заведом большое число (100), чтобы получить прямую зависимость с метаболической готовностью.
Физиологическое состояние бактерий, преобладающая среди членов бактериального блока экологическая стратегия и физиологическое разнообразие в гидролитическом бактериальном блоке различных почв сравнивали комплексным структурно-функциональным методом.
2.2. Результаты и обсуждение
2.2.1. Общая численность бактерий и динамика восстановления роста бактерий в процессе оттаивания образцов почв.
Как показали проведенные ранее исследования (Boyd et al., Moiroud a. Gounot, 1969, Horowitz etal., 1972, Friedmann, 1982, Cameron, et al., 1976, Vishniac, 1993.), разнообразие и численность бактериального сообщества антарктических почв, выявляемых, в том числе, и по числу культивируемых клеток, на несколько порядков ниже по своим показателям по сравнению с почвами иных регионов. Разница в показателях числа жизнеспособных культивируемых клеток может зависеть как от химического состава и генезиса исследованных образцов антарктических почв, так и от методов выделения жизнеспособных микроорганизмов.
Проведенное ранее изучение примитивных почв оазисов Ларсеманна Восточной Антарктиды показало, что общая численность бактерий в примитивных почвах влажных долин оазиса Ларсеманн не превысила 108 клеток в 1 г., это ниже значений, обычно регистрируемых в почвах умеренных широт. Также была выявлена высокая численность и доля жизнеспособных клеток в исследованных образцах (60 % и выше) свидетельствуют о высокой устойчивости бактериальных комплексов почв влажных долин к воздействию неблагоприятных факторов внешней среды в условиях Восточной Антарктиды.
В целом показатели численности, выявляемые по числу культивируемых клеток, из большинства исследованных нами образцов колеблются от 103 до 106кл/ г сух веса.
Полученные нами данные по числу культивируемых форм гетеротрофных эубактерий, показали, что их численность в целом сопоставима с численностью культивируемых форм гетеротрофных бактерий, выделяемых из ранее изученных образцов антарктических почв. Наиболее высокие показатели числа КОЕ были отмечены в образцах каменной мостовой (№ 5Д) с содержанием органики 8.8% - 2,3*![]()
![]() КОЕ на г и торфянистом горизонте (№14) с содержанием органики 8.74%. - 3,8*
КОЕ на г и торфянистом горизонте (№14) с содержанием органики 8.74%. - 3,8*![]()
![]() КОЕ на г.
КОЕ на г.
Таблица 2. «Численность бактерий, определяемая методом прямого счёта и число КОЕ на г на 5 сутки культивирования при комнатной температуре на TSA)».
№ Разреза и описание почвы | № образца | C, % | Прямой счёт | Число КОЕ на грамм на пятые сутки культивирования |
M3, мох | 1Д | 5,85 | 2.8* | 1,2* |
M3 почва в днище влажной долины, B 1-2 см | 3 | 0.85 | 2.8* | 4,9* |
M2, Bfungi, 1-2 | 3Д | 0,28 | 0.6* | Нет роста |
M4 0-7 песчаный мелкозём в наскальной ванне под сообществами эпилитных лишайников | 4Д | 3,3 | 2.2* | 1,8* |
NsM 10-20 O мерт. | 5Д | 8,8 | 3.8* | 2,3* |
10-15P1 каменная мостовая без признаков макробиоты | 10 | 0,08 | 4* | Нет роста |
10-06 с торфяным горизонтом | ||||
Km+V 0-2(4) см. Каменная мостовая со скоплениями одноклеточных водорослей | 13 | 0,12 | 2.9* | 5,3* |
T 2-4 см. Торфянистый горизонт, в основном мёртвая биомасса | 14 | 8,74 | 2.3* | 3,8* |
B1 4-10 см. Минеральный материал, ржаво-бурый крупнозернистый песок, есть признаки мицелия (грибов или актиномицетов) | 15 | 0,26 | 3.1* | 6,8* |
В целом, как видно из таблицы, наблюдается положительная корреляция между числом культивируемых клеток гетеротрофных бактерий и содержанием органического углерода, что отмечалось ранее в других образцах формирующихся почв данного региона. Полученные нами данные, согласуются с опубликованными ранее (Соина с соавт., 2012). Авторы делают предположение, что положительная корреляция числа колониеобразующих единиц гетеротрофных бактерий указывает на то, что значимая часть органического вещества лабильна и доступна для бактерий, но слабо связана с минеральной матрицей и слабогумифицирована. В целом число культивируемых клеток в исследованных образцах почв сопоставимо с такими же показателями а образцах древних мерзлых осадков Антарктиды.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
