
Рассмотрим составление сводки на примере однолетних самок. Поголовье самок на 20года принимается за исходную величину, т. е. за 100 %. 369 прохолостевших (непокрытых) самок составляют 30,4% (369*100:1212).
Вторую часть сводки составляют в следующем году (2001), От числа покрытых (принимая их за 100%), определяют % пропустовавших, абортировавших и НБР и др. (При необходимости эти виды потерь можно подсчитать по отдельности). Вычислять % этих потерь от числа всех самок, бывших на начало гона, не имеет смысла, т. к. непокрытые самки не могут пропустовать и тем более неблагополучно родить.
В категорию «всего самок без приплода» (ячейка Е12) включают прохолостевших, пропустовавших, абортировавших и НБР (Б12+Г12,+Д12), Благополучно ощенившими считают самок, у которых к моменту регистрации (45 дневный возраст щенков) сохранился хотя бы один живой щенок Цифру ячейки Ж12 можно также определить вычитанием из величины А12 ВеличиныЕ12.
Всех родившихся щенков, (ж.+м.), принимают за 100%. Количество ж.+м. щенков необходимо для вычисления средней по стаду плодовитости (количество ж.+м. щенков, полученных от благополучно родивших самок деленное на количество этих самок). Частой ошибкой является включение в количество щенков мертвых щенков от НБР самок. Этого делать не следует, потому что тем самым завышается плодовитость. Средняя по стаду плодовитость является очень важным показателем, характеризующим потенциал стада. % щенков, павших до регистрации вычисляют от живых.
Количество щенков, павших после регистрации в анализируемую сводку не включают, т. к. отход после регистрации не характеризует воспроизводительную способность стада, а зависит в основном от санитарного состояния фермы и качества кормов.
От однолетних самок в клеточных условиях получают очень мало щенков, и, казалось бы, не стоит по ним вести учет. Действительно, они не половозрелы, что выражается в большом количестве прохолостевших и пропустовавших самок (табл. 2), а также меньшей плодовитости. Однако характеристика репродуктивности однолеток дает представление о качестве подготовки молодняка к гону и, кроме того, известно, что покрытые однолетки в дальнейшем размножаются значительно лучше, чем не покрытые.
Главным видом потерь в соболеводстве является пропустование самок и потери в процессе совершенствования соболеводства не уменьшаются.
Если пропустование однолетних самок можно объяснить их половой незрелостью, то для пропустования более половины двухлеток и трети трехлеток (табл.2) такое объяснение вызывает сомнение. Тем более, что оставленные в основном стаде пропустовавшие, даже в четырех - и пяти - и шестилетнем возрасте по-прежнему не щенятся. Уменьшение количества пропустовавших в возрасте 4-10 лет (табл.1) объясняется выбраковкой всех не размножающихся в ранних возрастах. В хозяйствах, самок, не покрывшихся в двухлетнем возрасте и не щенившихся в трехлетнем возрасте экономически целесообразно выбраковывать. Как показывают данные таблицы 1, пропустование наступает при высокой плодовитости. Это наводит на мысль, что оно не связано с плохой подготовкой зверей к гону.
Таблица 2
Процент пропустования самок в хозяйствах России
Зверохозяйства | годы | Пропустовало самок | ||
Возраст | ||||
1 | 2 | 3 | ||
Салтыковский | 73,6 | 47,4 | 13,3 | |
Салтыковский | - | 40,4 | 25,0 | |
Салтыковский | 78,8 | 43,2 | 26,7 | |
Салтыковский | 80,8 | 48,6 | 26,1 | |
Салтыковский | 81,3 | 55,4 | 32,6 | |
Пушкинский | 80,9 | 65,2 | 64,8 | |
Пушкинский | 89,1 | 82,7 | 67,6 | |
Пушкинский | 72,8 | 48,5 | 33,5 | |
Пушкинский | 77,6 | 45,1 | 36,1 | |
Сомовский | 86,7 | 57,9 | 41,6 | |
Сомовский | 86,3 | 56,7 | 18,9 | |
Заря | 77,9 | 40,8 | 33,7 | |
Заря | 73,3 | 44,6 | 29,1 | |
Дикие | 39,4 | - | - |
В последнее время появились сообщения, свидетельствующие о несовпадении периода половой активности молодых самок с периодом соединения пар. Также имеются сведения, что при спаривании молодых самок с молодыми самцами, половая активность последних, недостаточна для полноценных покрытий. Плодовитость молодых самок, кроме однолеток, мало чем отличается от плодовитости взрослых.
Количество абортировавших и неблагополучно родивших самок в соболиных стадах обычно не велико. Даже меньше, чем у зверей других видов. Несколько большими показателями отличаются самки старше 10 лет и однолетки. У однолеток с возрастом этот показатель будет улучшаться, а у старых ухудшаться. В современных кормовых условиях самок старше 10 лет в стаде держать экономически не выгодно.
Для того чтобы составить полноценную сводку необходим правильно организованный первичный учет. Так сведения о воспроизводителности конкретной самки в течение жизни должны быть сосредоточены в одном месте. Это может быть трафаретка или племенная карточка. В соболеводстве, как ни в каком другом водстве, необходима компьютеризация племенного учета. В первичных записях не следует показатели гона отделять от результатов щенения. Это позволит выбрать правильное направление селекции для повышения репродуктивности стада.
Выводы:
1. Однолетние самки не являются половозрелыми, и их не следует включать в основное стадо.
2. Плодовитость молодых самок мало чем отличается от плодовитости взрослых.
3. Репродуктивность самок старше 10 лет резко снижается и их надо своевременно выбраковывать.
4. Пропустование одно-, двух-, и трехлетних самок, вероятно, в многих случаях связано с неправильной технологией гона.
Влияние БАВ на снятие теплового стресса у соболей
, ,
ФГБОУ ВПО МГАВМиБ им.
Аннотация. Показано влияние БАВ на снятие теплового стресса у соболей и воспроизводительную способность самок основного стада.
Ключевые слова: соболь, стресс, ПОЛ, БАВ.
Соболеводство - одна из немногих отраслей российского сельского хозяйства, которая исторически обладает определённым экспортным потенциалом и при правильной организации способна интегрироваться в мировую экономическую систему без чрезмерных капитальных вложений. Перед данной отраслью остро стоит проблема повышения естественной резистентности зверей в условиях производства, так как в ряде случаев используемая в хозяйствах технология не отвечает современным условиям, и звери вынуждены адаптироваться с большим напряжением физиологических систем. При этом во многих случаях происходит развитие стрессового состояния, и как следствие, снижается естественная резистентность организма, замедляется рост и развитие, снижается плодовитость, ухудшается качество шкурки, увеличивается заболеваемость и падеж.
Хищные пушные звери, как животные с хорошо развитой нервной системой, подвержены стрессу. Он возникает в результате резко изменяющихся внешних условий. Так, например, резкое изменение температуры, смена распорядка дня, переход от одного рациона к другому, длительное отсутствие воды и пищи, и другие внезапные воздействия вызывают стрессовое напряжение.[1]
Воздействие климатического стрессора на размножение соболя имеет ряд особенностей. В первую очередь, это наличие в годовом (фенологическом) цикле критических периодов, когда влияние этих факторов на показатели воспроизводства животных особенно ощутимо.
К ним относятся:
- период имплантации зародыша, начало истинной беременности (конец февраля - начало марта);
- истинная беременность и начальный период выкармливания молодняка (март-май);
- сезон спаривания - период, предшествующий гону, и собственно гон (июнь - июль, иногда август). [2,4]
Влияние метеорологических факторов на размножение соболей наиболее заметно и часто проявляется в отношении метеоусловий июня - июля. Это как бы узловая точка фенологического цикла, имеющая ведущее значение в определении будущей продуктивности животных. [5]
В этой связи интересно наблюдение, что при повышении температуры происходит накопление в крови активных форм кислорода, продуктов перекисного окисления липидов (ПОЛ), что свидетельствует о том, что значительное повышение среднесуточной температуры может вызывать у животных окислительный стресс. [6]
Поэтому вызывает интерес разработка и внедрение в соболеводство различных стресс-корректров, влияющих на стресс-лимитирующие структуры и системы организма. К таким средствам можно отнести тестируемый нами препарат, в состав которого входят такие биологически активные вещества как селенометионин, глутаминовая кислота, β-каротин, аскорбиновая кислота, витамин E и лецитин.
Селен – важнейший элемент антиоксидантной защиты организма. Он способен оказывать влияние на жизнедеятельность клетки как через изменение активности селеносодержащих белков, обладающих антиоксидантными свойствами, так и через редокс-регуляцию множества метаболических функций клетки. Селен является компонентом фермента глутатионпероксидазы, которая вместе с пептидом глутатионом защищает клетки от разрушающего воздействия перекиси водорода.
Метионин является донором метильной группы, необходим для синтеза: холина, фосфолипидов. Снижает риск токсических поражений и жировой инфильтрации печени. [3]
Глутаминовая кислота занимает центральное место в метаболизме свободных аминокислот. Кроме того, глутаминовая кислота играет исключительно важную роль в обезвреживании образующегося в тканях аммиака. При воздействии стрессовых факторов отмечено стимулирующее влияние глутаминовой кислоты на дыхание и окислительное фосфорилирование в митохондриях печени и нормализацию метаболизма в ней.
Неферментативную антиоксидантную защиту организма обеспечивают витамины С, Е и β-каротин, которые инактивируют на разных уровнях высоко токсичные формы кислорода, непрерывно образующихся в процессе нормальной жизнедеятельности любой клетки.
Лецитин является основным питанием всей нервной системы (входит в состав оболочек нервных волокон; обеспечивает нормальный обмен фосфолипидов), снижает концентрацию жирных кислот в крови, улучшает функцию печени и почек, помогает усвоению жирорастворимых витаминов A, D, E и K.
Целью наших исследований является изучение влияния указанных выше веществ на биохимические показатели крови соболей при тепловом стрессе.
Исследования проводились на базе «Пушкинский» Пушкинского района Московской области, на кафедре органической и биологической химии МГАВМиБ им. в гг. При проведении научно-производственных опытов использовали самок-однолеток черного соболя.
По принципу аналогов нами были сформированы две группы самок-однолеток по 20 голов в каждой, средней массой 1.1 кг.
1 группа – контрольная – дача препарата не осуществлялась, соболя подвергались тепловому стрессу.
2 группа – опытная – препарат задавали с кормом в терапевтической дозе.
Взятие крови производилось из капилляров когтя до воздействия стрессового фактора (норма), перед дачей препарата при воздействии стрессового фактора и на 5 сутки после дачи препарата. Биоматериалом для биохимических исследований служила сыворотка крови. Содержание в сыворотке общих липидов определяли с фосфованилиновым реактивом и H2SO4. Концентрацию малонового диальдегида определяли с помощью тиобарбитуровой кислоты. Определение диеновых конъюгатов и диенкетонов проводили модифицированным методом Плацера.
Результаты наших исследований показали, что биологически активные вещества препарата ведут к снижению уровня перекисного окисления липидов и положительно влияют на биохимические показатели крови зверей.
Ярко выраженное повышение среднемесячной температуры июля на 45% (с +18 до +26,1оС) по сравнению с нормой для данного региона и повышение среднесуточной температура до 30 оС и выше (рис. 1.) послужило причиной возникновения окислительного стресса у соболей. При этом в крови животных резко возросли концентрации общих липидов и продуктов ПОЛ.
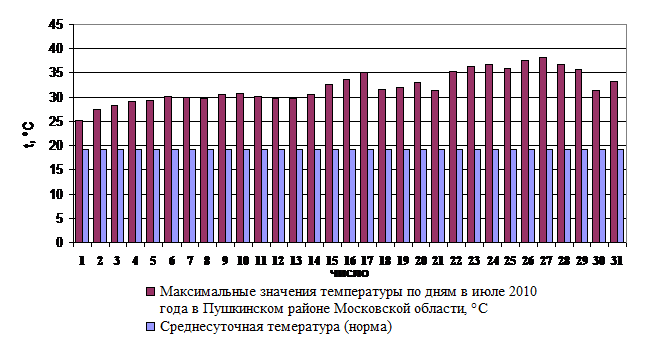
Рис. 1. Динамика изменения температуры на июль 2010 г.
Активные вещества испытываемого препарата снизили содержание общих липидов в сыворотке крови соболей опытной группы на 14,94% по сравнению с контрольной (рис. 2).
Более высокое содержание общих липидов в контрольной группе коррелирует с высоким содержанием продуктов перекисного окисления липидов в этой группе.
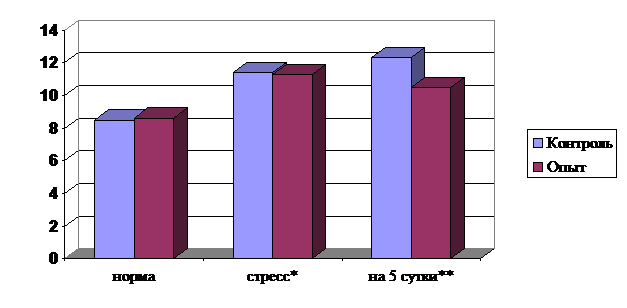 Рис. 2. Влияние препарата на содержание общих липидов в плазме крови у соболей (г/л)
Рис. 2. Влияние препарата на содержание общих липидов в плазме крови у соболей (г/л)
Примечание: * - дача препарата при воздействии стрессового фактора,
** - изменение показателей на 5 сутки после дачи препарата.
Содержание ТБК-активных продуктов оценивали по количеству малонового диальдегида (МДА) – вторичного продукта перекисного окисления липидов, который, являясь мутагеном и цитотоксином, способен ингибировать синтез белка, нуклеиновых кислот и т. д. В сыворотке крови опытной группы содержание МДА было меньше по сравнению с контролем на 40.27% (рис. 3), что свидетельствует о снижении токсичности сыворотки крови.
Количество первичных продуктов ПОЛ – диеновых конъюгатов в сыворотке крови соболей опытной группы было ниже относительно контрольной группы на 23.45%, в которой происходит гиперактивация процессов ПОЛ (рис. 4).
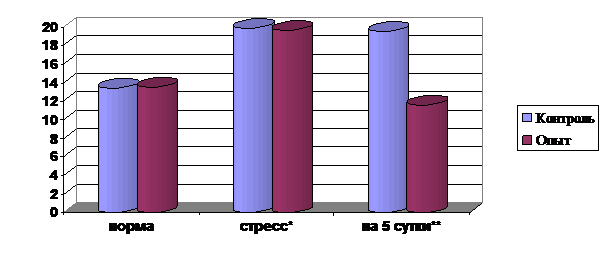
Рис. 3. Влияние препарата на показатели ТБК-активных продуктов (малоновый диальдегид) в сыворотке крови соболей (мкмоль/л)
Примечание: * - дача препарата при воздействии стрессового фактора,
** - изменение показателей на 5 сутки после дачи препарата.
Содержание вторичных продуктов ПОЛ – диенкетонов в сыворотке крови в опытной группе было ниже на 41.67% по сравнению с контрольной группой, в которой наблюдается пролонгация процессов переокисления с повышенным образованием карбонильных продуктов ПОЛ (рис. 4).
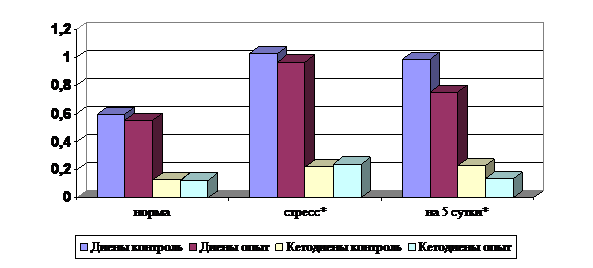
Рис. 4. Содержание диеновых конъюгатов и диенкетонов в сыворотке крови у соболей (едА/мл)
Примечание: * - дача препарата при воздействии стрессового фактора,
** - изменение показателей на 5 сутки после дачи препарата.
После окончания гона были получены следующие результаты. В контрольной группе в охоту пришли 70% самок, из них покрылось 21,5%. В опытной группе в охоту пришли 80% самок, однако число покрывшихся самок составило 37,5%.
Таким образом применение препарата во время гона позволило получить на 0,41 щенка больше на самку в опытной группе.
Можно заключить, что физиолого-биохимическая оценка гомеостаза у экспериментальных животных подтвердила у соболей в период повышения среднесуточной температуры наличие окислительного стресса и увеличение содержания в крови общих липидов в контрольной и опытной группах на 34,2% и 30,8% соответственно. Содержание продуктов ПОЛ в контрольной и опытной группах также увеличилось на: 47,3% и 45,2% для МДА, 72,7% и 76,0% для диеновых конъюгатов, 71,9% и 91,1% для диенкетонов.
Активные вещества препарата повышают активность антиоксидантной защиты организма, достоверно снижая уровень ПОЛ, положительно воздействуя на биохимические показатели крови у животных опытной группы. Так, показатели общих липидов снизились на 14,9%, МДА - на 40,3%, диеновых конъюгатов и кетодиенов на 23,5% и 41,7% соответственно, по сравнению с контролем.
Препарат может быть рекомендован для профилактики и коррекции окислительного стресса у соболей в условиях производства.
Литература
1. Балакирев / , – М.: КолосС, 2006, 344 с.
2. Белов , погода и размножение соболя полуострова Камчатка / , // Промысловые звери РСФСР (пространственные и временные изменения населения): Сб. науч. тр. ЦНИЛ Главохоты РСФСР. - М., 1982.-с. 179-190.
3. Зенков стресс. Биохимический и патофизиологический аспекты / , , - М.: Слово, 2001, 556 с.
4. Кукушкин М, А. Некоторые механизмы экзогенной регуляции репродуктивного процесса в популяциях соболя // Кукушкин основы охраны и воспроизводства охотничьих ресурсов. - М., 1987, гл. 7. - с. 73-81.
5. Кукушкин содержания и особенности влияния метеорологических факторов на размножение соболя в начальный период искусственного разведения (1932—1950 гг.) / // Первое Всесоюзн. совещ. по проблемам зоокультуры: Тезисы докл. - М., 1986. ч. 1. с. 227-229.
6. , Эммануэль свободнорадикальных реакций окисления в молекулярных механизмах старения живых организмов // Успехи химии. – 1983. –Т. 52. –с. 353-372.
ДНК-маркеры: новые возможности в практике
разведения соболя
, кандидат биологических наук
, .кандидат биологических наук
Учреждение Российской академии наук Институт общей генетики им. РАН
Учреждение Российской академии наук Институт биологии развития им. РАН
Сохранение природных популяций соболя, вновь созданных пород и типов этого вида является одной из актуальных проблем на сегодня. Природные популяции соболя, распространенные от Приуралья до Дальнего Востока, географически разделены на региональные типы, которые обладают рядом морфологических особенностей. За последние 300 лет популяции соболя претерпели в результате неорганизованного промысла и природных катастроф значительные изменения численности, причем некоторые из них исчезли. Общий ареал вида к концу 19-го века распался на ряд отдельных очагов. На государственном уровне был принят ряд мер: создание новых соболиных заповедников – Печоро-Илычский (1930 г.), Кондо-Сосьвинский (1929 г.), Алтайский (1932 г.), Саянский (1939 г.), Сихотэ-Алинский (1935 г.) и Кроноцкий ( 1934 г.), и расширению возможностей ранее созданных (Баргузинский, 1916 г.); создание заказников, например, в Бурятии в разное годы существовало от 3 до 22 заказников. Дополнительно к этому были введены запреты на добычу соболя, которые в отдельных районах носили длительный характер. Кроме того, были предприняты беспрецедентные по своему масштабу меры по расселению соболя на огромных территориях, включая даже те, где он ранее не встречался. Общая численность реинтродукцированных соболей за весь период работ по восстановлению его ареала от Урала до Дальнего Востока составила около 20 тыс. особей. В качестве примера можно привести динамику численности соболя в Новосибирской области. Здесь, в результате принятых мер по расселению соболя в Западной Сибири стали возможными естественные миграции на территорию Новосибирской области из сопредельных регионов, в которых проводилась реинтродукция соболя. Динамика численности показывает, что в этой области ареал соболя в 1696 году составлял 6.3 млн. га (данные по чертежам Сибирских городов и земель, составленных известным сибирским картографом Семеном Ульяновичем Ремезовым). В 1913 году эта территория уменьшилась в 4 раза, а в 20-ых годах прошлого века соболь в Новосибирской области исчез полностью. Новое заселение соболем этих территорий отмечено только в 1962 году, и началось оно на границе с Томской областью, куда соболь проник в результате естественных миграций из таежных угодий бассейнов рек Васюгана и Парабели. К настоящему времени соболь расселился в таежной зоне этой области повсеместно, отмечены заходы зверьков даже в лесостепные районы. Ареал соболя в области стабилизировался. Далее, восстановленный ареал соболя Западной Сибири постепенно слился с его ареалом на северных отрогах Саян и Кузнецкого Алатау, а на севере, по долине Оби и ее притоков, расширился практически до Обской губы. Анализ миграционных путей показал, что соболя из сравнительно хорошо сохранившихся очагов Восточной Сибири перевозили в Западную Сибирь, регионы Крайнего северо-востока и другие. На этой основе популяции соболя Восточной Сибири названы популяциями-донорами.
В этот же период после того, как были отлажены методы отлова и транспортировки соболя, были предприняты попытки по его разведению. Это перспективное направление сельскохозяйственного животноводства может оказать значительное влияние на сохранение природных популяций. Все эти меры по восстановлению популяций соболя внесли свои коррективы в генофонд этого вида. Глобальное изменение условий жизни соболя возможно при использовании всей генетической изменчивости вида, которая служит основой эволюции, его способности адаптироваться к меняющимся условиям внешней среды, в том числе и к условиям клеточного содержания. Об уровне изменчивости природных популяций соболя можно судить по примеру результатов сравнительного анализа площади шкурки из популяций Амура, п-ва Камчатка, Якутии и Урала: выявлены значительные различия между популяциями (площадь различается в два раза). Далее эти исследования были расширены за счет привлечения молекулярно-генетических методов. Исследования проводились с инвентаризацией геномов природных популяций от Урала до п-ва Камчатка (места взятия выборок: хребет Хамар-Дабан, Енисейский бассейн, Ленский бассейн, Приуралье, Восточные Саяны, правобережный Амур и п-в Камчатка). На базе этих популяций были сформированы клеточные. Задачами исследования являлись: выявление особенностей генофондов природных и клеточных популяций, проведение сравнительного анализа, который позволил выявить изменения генофонда соболя после 80 лет селекции. Для оценки результатов селекции и формирования современных подходов к отбору производителей собран банк ДНК соболя (2 тыс. образцов). Впервые для этого вида отработаны маркеры ядерного и митохондриального геномов. Проведен сравнительный молекулярно-генетический анализ природных и клеточных популяций с использованием этих маркеров (D-петля, 12 микросателлитных локусов и три ISSR-маркера).
Анализ митохондриального генома был выполнен на наиболее вариабельной области мтДНК животных – последовательности регуляторного района (D-петля). Исследованный фрагмент находится в области, примыкающей к гену тРНK-Pro, и включает точку терминации репликации D-петли. Для характеристики генетической изменчивости клеточного соболя проведено секвенирование фрагмента D-петли. Были получены фрагменты длиной 426 н. п., соответствующие области с 15486 по 15912 нуклеотид в полном митохондриальном геноме соболя. Показано, что соболь обладает значительной митохондриальной изменчивостью. Всего определено 117 последовательностей D-петли клеточного соболя, которые образуют 34 гаплотипа. В анализируемом фрагменте выявлена высокополиморфная область с 1 по 161 нуклеотид и консервативная, в которой не найдено мутаций, за исключением небольшого поли-G-блока в районе нуклеотидов.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
