

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
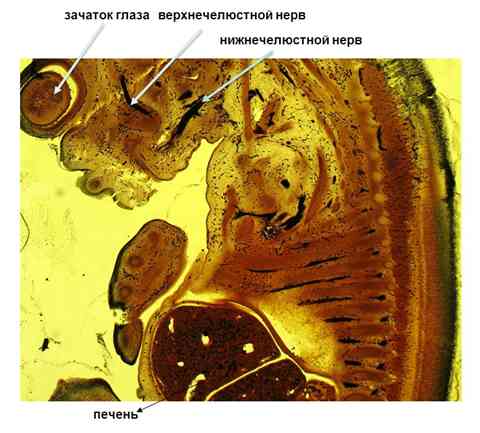
Рис. 1 сагиттальный срез эмбриона 17 ст. Импрегнация по Бильшовскому-Буке. Об. 10. Ок.10
В дальнейшем главные ветви узла тройничного нерва приобретают пучковидное строение сначала в местах отхождения от узла, а затем на всем протяжении. К этому времени вторичные ветви состоят из прямых нервных волокон, формирующих компактный ствол. Наконец, в составе основных ветвей тройничного нерва нарастает количество пучков, и вторичные ветви также приобретают пучковидное строение.
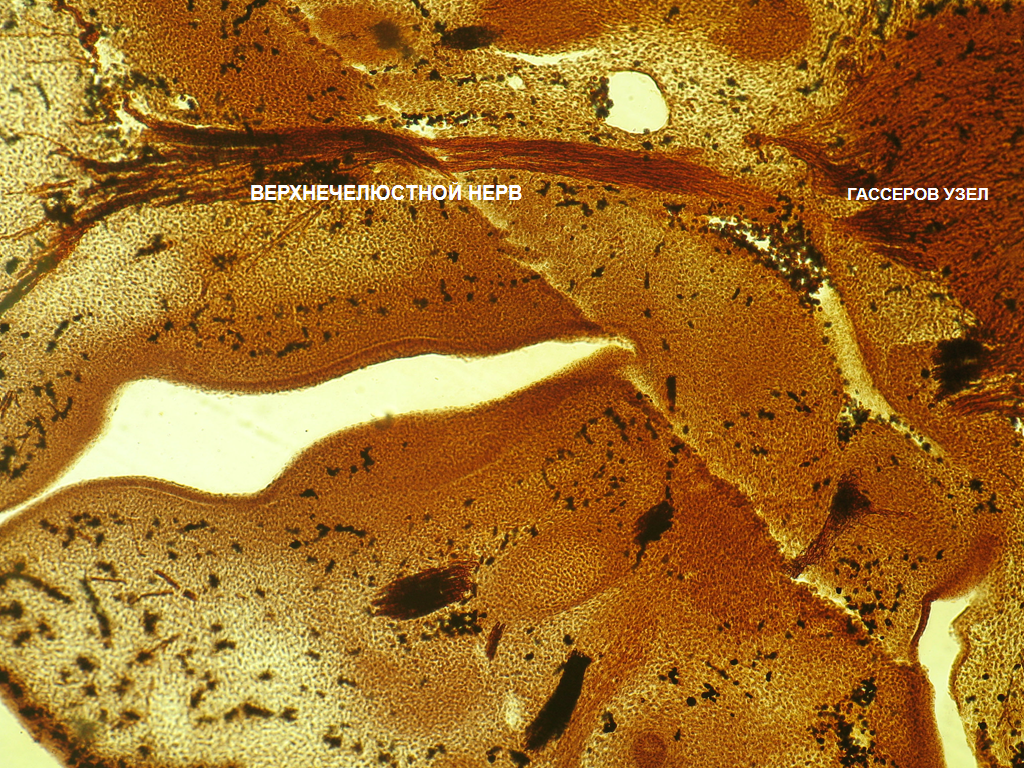
Рис. 2 - срез эмбриона 18 ст. Импрегнация по Бильшовскому-Буке. Об. 10. Ок.10
Процесс развития и дифференцировки ветвей тройничного нерва распространяется от центра (от узла) к периферии.
На протяжении исследованного периода развития эмбрионов человека в построении ветвей тройничного нерва можно выделить 4 стадии.
На первой стадии ветви тройничного нерва состоят из сильно извитых, переплетающихся между собой нервных волокон; на второй стадии ветви тройничного нерва представлены рыхлым пучком и формируют вторичные нервные стволы и закладки двигательных ветвей; на третьей стадии ветви тройничного нерва представлены компактными стволами, формируют все вторичные ветвления, характерные для дефинитивного состояния; на четвертой стадии происходит удлинение ветвей, нарастает количество конечных ветвлений чувствительных и двигательных нервов и начинает выявляться пучковидное строение основных стволов тройничного нерва.
Флуоресцентно - гистохимическое исследование
Единичные холинергические нейроны со слабовыраженной реакцией на сульфид меди впервые выявлялись у 8-и недельных эмбрионов. Такие нейроны, в количестве 8-11 на одном срезе, располагались в центре и на периферии тройничного ганглия.
Также, единичные холинэстераза-положительные нейроны встречались по ходу нервных волокон растущих ветвей n. glossoрharyngеus, n. maxillaries, n. mandibularis, по ходу нервных волокон лицевого нерва, в месте отхождения большого каменистого нерва и по ходу барабанной струны, узле коленца и близлежащих узлах статически-слухового нерва, по ходу нервных волокон растущих ветвей блуждающего нерва, но в значительно большей степени они выявлялись на стенке пищевода в составе пищеводного сплетения, в шейных спинномозговых узлах.
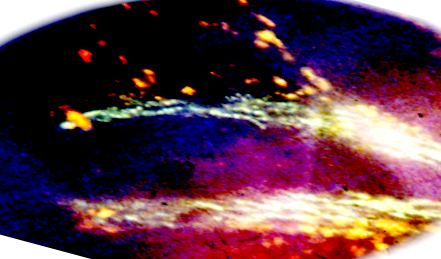
Рис. 3 - Флуоресценция катехоламинов (зеленое) и серотонина (желтый) вдоль небной артерии. Стадия 23. Инкубация в глиоксиловой кислоте. Об. 10. Ок. 10
Исследование срезов с инкубацией в глиоксиловой кислоте для выявления катехоламинов показало появление единичных интенсивно флюоресцирующих нейронов (SIF cells) у 10-и недельных эмбрионов. Единичные SIF-нейроны располагались вблизи скоплений нейробластов, формирующих каудальный и краниальный ганглии n. glossoрharyngеus и n. vagus, а также по ходу их ветвей, вблизи формирующихся последовательно ganglion ciliare, ganglion pterygopalatinum и ganglion oticum, а также по ходу ветвей тройничного нерва, вблизи группы нейронов, формирующих узел коленца, рядом со скоплениями нейробластов, формирующими каудальный и краниальный ганглии блуждающего и языкоглоточного нервов, а также по ходу их ветвей, возле скоплений симпатобластов, формирующих ganglion axillar superior, затем ganglion vertebrale и ganglion stellatum.
Между краниальными ганглиями блуждающего и языкоглоточного нервов, а также вблизи Гассерова узла, вокруг всех ганглиев pars cervicalis truncus symphaticus во всех случаях наблюдались 1-2 мелких параганглия содержащие клетки со слабой специфической флюоресценцией цитоплазмы и нелюминесцирующими ядрами.
Также, в большинстве случаев наблюдений, по ходу сино-каротидной ветви n. glossoрharyngеus, по ходу n. ophtalmicus, n. maxillaris и n. mandibularis, по ходу ветвей лицевого нерва, чаще в местах ветвления их, по ходу межузловых ветвей шейного отдела симпатического ствола выявлялась группа нейронов, выказывавших слабую специфическую флюоресценцию катехоламинов.
На последовательных стадиях развития челюстно-лицевого аппарата люминесцентно-гистохимическими методами исследования на 16 стадии эмбриогенеза выявляются люминесцирующие кровеносные сосуды, а затем, на 19 стадии, в них определяются нервные волокна, содержащие ацетилхолин, позднее, на 20 стадии появляются серотонинсодержащие, а на 22 стадии – катехоламинсодержащие клетки. Появление первичных кровеносных сосудов в ЧЛО с большой площадью ветвления совпадает с образованием первичного неба. В местах расположения медиаторспецифичных нервных окончаний наблюдаются участки дифференцировки мезенхимы.
Наибольшее содержание серотониновых рецепторов отмечается на 22 стадии развития, что предшествует образованию вторичного неба.
Появление SIF-клеток совпадает с ускорением процессов гистогенеза и дифференцировки клеток области рта и носа.
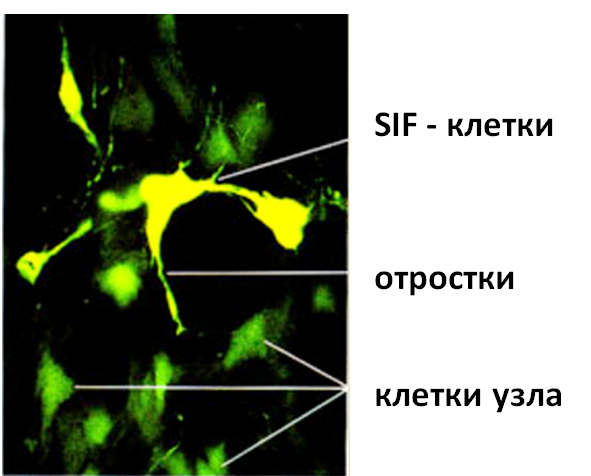
Рис. 4 -. Тройничный ганглий. Метод Фалька-Хилларпа. Об.40. Ок.10. Изображение обработано в ПО «Морфотест»
Таким образом, имеющаяся связь по содержанию нейромедиаторов в мезенхимальных структурах развивающегося ЧЛО, а также в медиаторспецифичных нервных окончаниях может быть в основе одного из важных механизмов роста и дифференцировки клеток в процессе формирования вторичного неба. Мы предполагаем, что исследуемые нейромедиаторы могут являться индукторами дифференцировки тканей в процессе раннего пренатального развития.
Влияние дефицита нейротрансмиттеров крови матери на развитие ЧЛО эмбриона
Через 1 сутки после экспериментального воздействия, состоявшего в блокировании нейромедиатора (независимо от его вида) в крови беременной самки крысы, у плода в разных отделах развивающегося тройничного ганглия регистрируется гибель нейральных клеток. Группы апоптотических телец обнаруживаются в тройничном узле, а также по ходу верхнечелюстного нерва и артерии. Достаточно часто погибшие клетки можно встретить в просвете верхнечелюстной артерии, куда выводятся остатки погибших клеток из нейроэпителиального пласта.
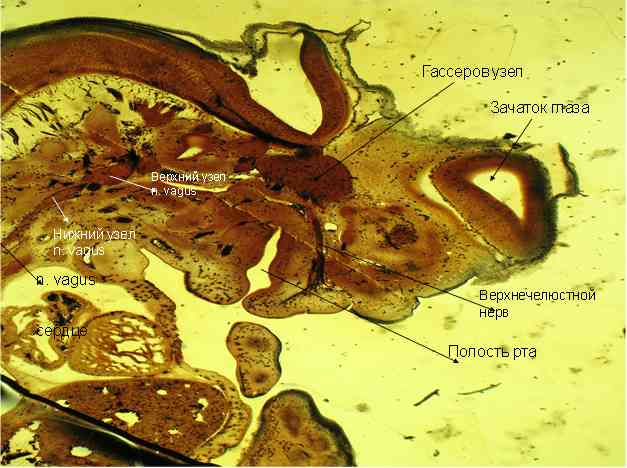
Рис.5 - Срез ЧЛО эмбриона крысы. Импрегнация по Бильшовскому-Буке. Об. 10. Ок.10
Изменения нервного аппарата при моделировании дефицита ацетилхолина
У крыс группы сравнения нервные элементы ЧЛО были представлены небольшим ганглиозным аппаратом и единичными нейронами с многочисленными нервными волокнами, распространяющимися от тройничного узла в сторону тканей-мишеней и теряющихся в этих тканях.
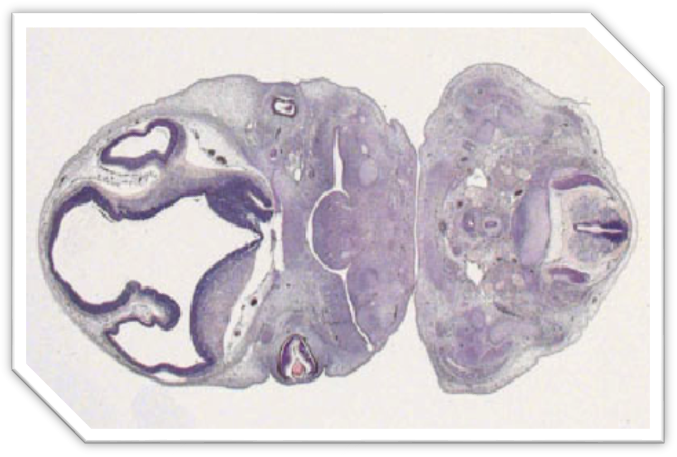
Рис. 6 – Срез эмбриона крысы. Окраска по , Об. 20. Ок.10
В динамике эксперимента у крыс наблюдали выраженные изменения в нервном аппарате, которые имели тенденцию к нарастанию. Значительно увеличивалось количество ганглионарных нейронов, находящихся в состоянии дистрофии и атрофии. Наиболее измененными оказались нейрон-содержащие периартериальные области, в несколько меньшей другие. Среди скоплений ганглиозных клеток выявлялись истинные явления нейронофагии и сателлитоз. Неравномерность хода нервных элементов нарастала. Среди нейроцитов обнаруживались также клетки с пикнозом ядер, вакуолизацией цитоплазмы. Единичные ганглионарные клетки подвергались невролизу.
Морфометрические исследования подтвердили указанные положения. Максимальная плотность нейронов выявлялась в периартериальной и межчелюстной областях, и там же отмечались максимальные изменения этого показателя при нарастании срока. Размеры ядер нейронов зависели от локализации в несколько меньшей степени, но варьировали по тем же закономерностям. В максимальной степени уменьшалась средняя площадь сечения ядер ганглионарных клеток, расположенных в периартериальной и межчелюстной областях (все Р<0,01). В периартериальной области и, в меньшей степени, венозной области плотность нервных элементов при развитии дефицита медиатора достоверно увеличивалась, а в межчелюстной области - прогрессивно уменьшалась (все Р<0,01). Увеличение плотности обычно сопровождалось некоторым утолщением нервных волокон.
Таким образом, развитие состояния дефицита нейромедиатора сопровождалось характерными изменениями вегетативных элементов ЧЛО, которые затрагивали как тела ганглионарных клеток, так и проводящие нервные элементы.
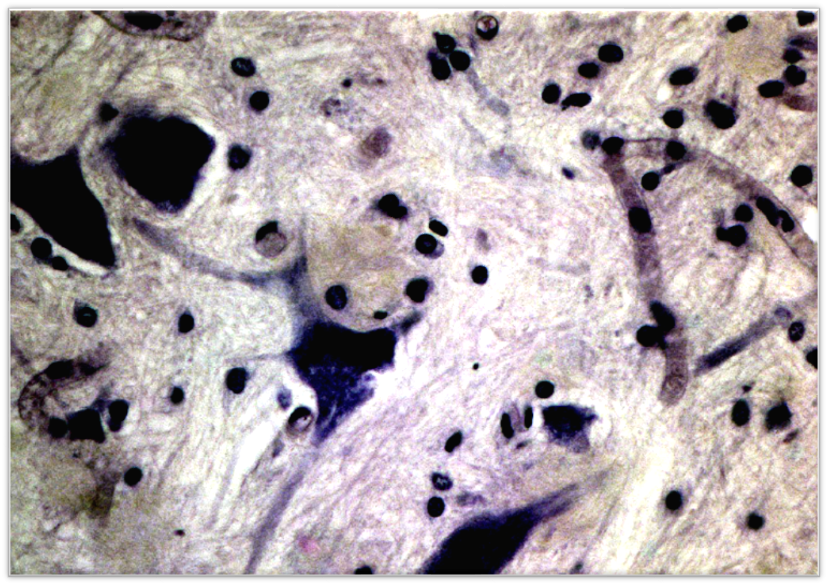
Рис.7 – Клетки тройничного узла эмбрионов основной группы. Окр. толуидин. Об. 40. Ок. 10
При исследовании под большим увеличением были обнаружены более дифференцированные изменения нейроцитов и нервных волокон в ткани ЧЛО. Большинство нейронов находилось в состоянии острого раздражения или дегенерации вплоть до нейролиза с формированием «клеток-теней». На этом фоне отчетливо было заметно появление гиперхромных нейроцитов с большим количеством хорошо окрашенных отростков, что свидетельствовало об их функциональной активности. Имелись также единичные двуядерные нейроны. Сателлитоз не был столь резко выраженным.
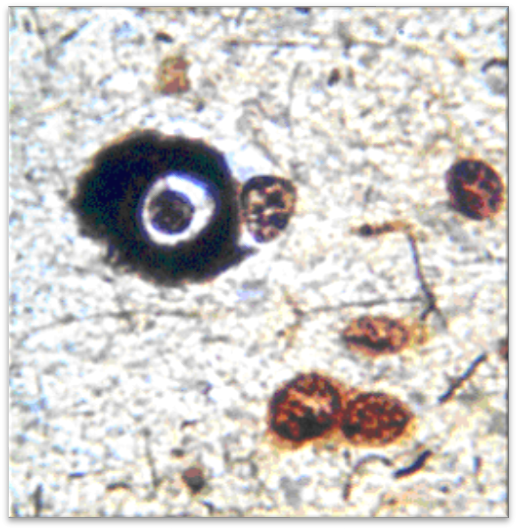
Рис. 8 – Клетки узла на 16 день. Серебрение. Об.40. Ок.10
При окраске по Нисслю и серебрении подтверждалась выраженная мозаичность поражения нейронов. Гиперхроматоз отдельных нейронов сочетался с дистрофией и нейролизом других клеток. Часть нейронов имела вид клеток-теней.
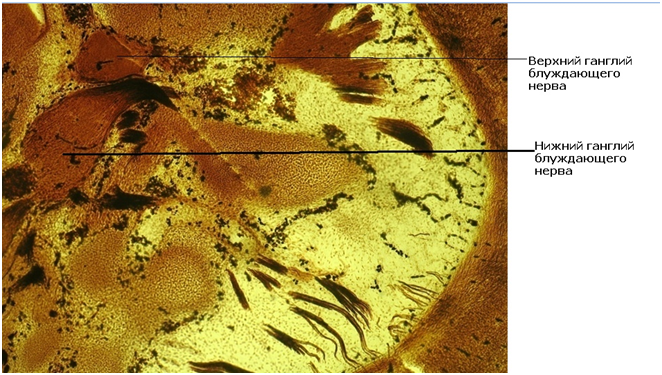
Рис.9 – Срез головного конца эмбриона основной группы. Окр. Бильшовский-Буке. Об. 10. Ок. 10
Характерной была мозаичная аргентофилия нейронов, набухание и извитость нервных волокон.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
