

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Густота ветвления нервных волокон и форма окончаний изменилась: вместо равномерного трехмерного рисунка с отчетливыми слоями нервные элементы местами исчезали или образовывали искривленные короткие кустикообразные скопления, а в отдельных местах - образовывали плотные ориентированные скопления, направленные вглубь (рис. 10).
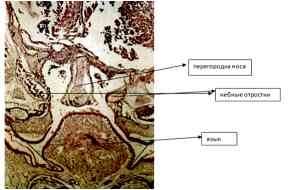
Рис. 10 – Срез ЧЛО эмбриона основной группы (16 день). Окр. H&E. О. 10. Ок. 10
Проявления отклонений в развитии неба были наиболее заметны в передней его части (рис. 11). В области первичного неба и передней части твердого неба нос был укорочен. Вертикальная часть его хрящевой перегородки отсутствовала. Двусторонние расщелины обнаруживаются в области первичного среднего небного треугольника, а перегородка носа отклоняется в сторону более узкой расщелины. В передней части твердого неба, боковые отростки неба очень малы или имеют неправильную конфигурацию. В задней области у нескольких животных были найдены аномалии в виде щитовидно-язычной щели. Кроме того, уменьшился объем хрящевой носовой капсулы в средней части твердого неба. Само небо выглядит нормальным, начиная с середины твердого неба назад.
У эмбрионов крыс, подвергшихся экспериментальному воздействию с блокированием ацетилхолина на 13 сутки эмбрионального развития, в области первичного неба хрящевая перегородка носа и носовая капсула были хорошо разграничены. Боковые небные отростки в этой области горизонтальны и сращены с носовой перегородкой. Во всех остальных участках неба небные отростки направлены в вертикальном направлении вдоль языка.
Таким образом, дефицит ацетилхолина на ранней постимплантационной стадии (Е9) оказывает повреждающее действие на развитие исследованных отделов и оставляет длительный и, видимо, необратимый след в их структурной организации.
Изменения нервного аппарата ЧЛО при деплеции норадреналина
При дефиците норадреналина у эмбрионов крыс первой группы на 14 сутки обнаруживали набухание части нейронов, явления их дистрофии и частичный невролиз. Общее количество волокон увеличивалось, они утолщались, но равномерность их расположения в ткани нарушалась: наряду с участками относительно обедненными появлялись плотные скопления нервных волокон. В последующих стадиях в скоплениях нейронов на фоне нарастания явлений дистрофии нейроцитов выявлялись также гипертрофированные клетки с большим ядром, в ряде случаев - двуядерные. Скопления нейронов вокруг сосудов представляли пеструю картину дистрофированных и погибающих нейронов и ганглионарных клеток в состоянии гипертрофии. Нервные волокна были распределены неравномерно, становилось очевидным преимущественное сохранение и уплотнение сети волокон, распространяющихся периваскулярно.
По данным морфометрии, максимальная плотность нейронов выявлялась в периартериальной области, наименьшая — в области носовой перегородки. Аналогичная закономерность была характерна для размеров ядер этих нейронов и плотности окружающих нервных волокон.
Развитие состояния дефицита норадреналина сопровождалось уменьшением плотности нейронов в скоплениях при возрастании общей плотности нервных окончаний. При дефиците норадреналина во всех изучаемых отделах, начиная с 15 сут, достоверно увеличивалась средняя площадь сечения нервных волокон.
Таким образом, развитие состояния дефицита норадреналина сопровождалось характерными изменениями вегетативных элементов, которые затрагивали как тела нейроцитов, так и проводящие нервные элементы области.
Нервные волокна, проходящие в составе нервных пучков в мезенхиме неба, интенсивно окрашивались серебром, местами обнаруживались варикозные вздутия. В то же время заметных изменений нервных проводников, расположенных в строме, не отмечалось. Нейроны микроганглиев неравномерно импрегнировались серебром. Их количество практически не изменялось, но картина становилась более пестрой: на фоне уменьшенных в объеме клеток с пикнотическим ядром и клеток-«теней», в которых не выявлялись ядра и ядрышки, присутствовали гипертрофированные клетки с аргентофильной цитоплазмой и большим количеством хорошо окрашенных отростков. Это свидетельствовало о различной степени выраженности функциональной активности нейронов
Неравномерность плотности нервных волокон, идущих по ходу небных отростков и крупных сосудов, была отчетливо видна, волокна в области скоплений были утолщены. В участках, расположенных на границе максимально токсически измененных очагов и малоизмененной ткани, ход афферентных и эфферентных нервных проводников прослеживался более отчетливо.
При исследовании фронтальных срезов ЧЛО эмбрионов крыс, подвергнутых эксперименту с ингибированием синтеза норадреналина, аномалии замечены по всей длине неба. Нос укорочен и заметно отклоняется в сторону от расщелины. Хрящевая носовая перегородка раздвоена по всей длине. Небные отростки расположены горизонтально, выше языка. Формирование кости заметно отстает по сравнению с контролем.
Изменения нервного аппарата ЧЛО при деплеции серотонина
При моделировании дефицита серотонина у крыс наиболее частыми проявлениями морфологических изменений в нейроцитах нервных сплетений являлись набухание и вакуолизация их цитоплазмы. Наибольшие изменения нервного аппарата выявлены в тройничном узле, где к 14 сут значительно увеличивалось количество дистрофически измененных нейроцитов, возрастало число атрофированных нейроцитов. Среди нейроцитов обнаруживались также клетки с пикнозом ядер, вакуолизацией цитоплазмы. В небном сплетении определялись также различные изменения нейронов от умеренной дистрофии или атрофии до сморщивания, острого набухания, часто с отложением в нервных клетках липидов типа липофусцина. Встречались гипертрофированные и двуядерные нервные клетки. Изменения нервных волокон и нервных проводников характеризовались их набуханием, гомогенизацией, избыточной аргирофилией и фрагментацией. С течением времени изменения приобретали еще более выраженный характер, хорошо была заметна мозаичность состояния нейронов: атрофированные нейроны, клетки-«тени» чередовались с гипертрофированными аргирофильными клетками, имеющими разветвленную сеть волокон вблизи перикариона.
Морфометрическое исследование количественно подтвердило характер описанных изменений. Развитие дефицита серотонина сопровождалось уменьшением плотности нейронов и проводящих элементов во всех изучаемых структурах, но в особенности в тройничном узле. Параллельно уменьшались средние размеры ядер нейронов, и возрастала толщина волокон по профилю поперечного сечения.
Структурные изменения в узлах характеризовались сочетанием дистрофических и регенераторных процессов. Дистрофические, атрофические изменения тел и отростков чувствительных нейронов развивались параллельно с гипертрофией части нейронов. В патологический процесс, наряду с изменениями тел и отростков нейронов, вовлекались также нервные волокна, проходящие в ганглиях и направляющиеся к головному мозгу (рис. 14). В отношении нервных волокон было характерно общее уменьшение их числа при некотором увеличении толщины и извитости.
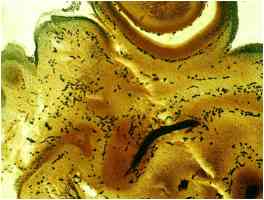
Рис. 11 – Срез ЧЛО эмбриона крысы. Окр. Бильшовский-Буке. Об. 10. Ок. 10.
Можно было наблюдать набухание, варикозные утолщения, вакуолизацию, а также зернисто-глыбчатый распад преимущественно мякотных нервных волокон. При этом тонкие безмякотные нервные волокна, как правило, страдали в меньшей степени. Появлялись короткие прямые, как бы оборванные, окончания.
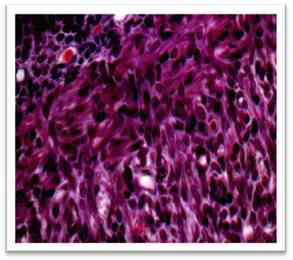
Рис. 12 – Срез крылонебного узла эмбриона крысы. Окр. фуксин. Об. 40. Ок. 10
Морфофункциональные изменения в развивающемся небе
У животных наблюдается двусторонняя расщелина, которая частично продолжается в твердое небо в виде расщепления по средней линии. Нос сильно притуплен и имеет раздвоение хрящевой носовой перегородки. Во всех областях расщелины, формирование кости выглядит неполноценным. Несмотря на двусторонний характер расщелины, носовая перегородка отклоняется в одну сторону. В области середины твердого неба, несмотря на то, что боковые нёбные отростки срослись и сформированы нормально, перегородка носа раздвоена.
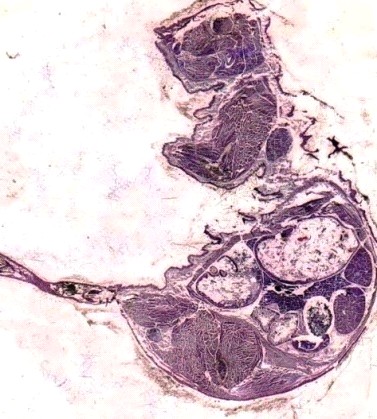

Рис.13. Срез ЧЛО эмбриона крысы первой группы. 16 день. Окр. H&E. Об. 10. Ок. 10
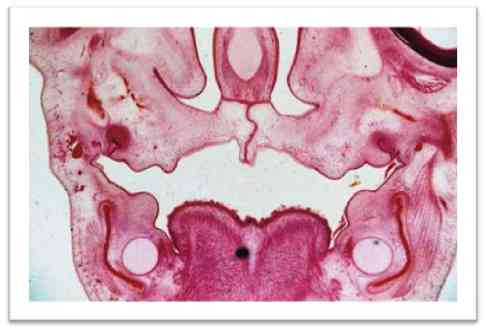
Рис.14. Срез ЧЛО эмбриона крысы второй группы. 16 день. Окр. H&E. Об. 10. Ок. 10
Гистохимическое исследование
В результате исследования, описанного ранее, нами были установлены сроки и закономерности становления медиаторного этапа челюстно-лицевой области, выражением чего стало появление медиаторспецифичных нервных окончаний и дальнейшее нарастание их плотности. Сопоставив сроки и фазы эмбрионального развития крысы и человека, мы адаптировали полученные данные к экспериментальному исследованию, которое описывается в данной главе.
Таким образом, холинергические нервные окончания должны появляться в ЧЛО примерно на 13 сутки эмбрионального периода. Адренергические – на 13,5 сутки.
Экспрессия серотонина на 16 сутки эмбриогенеза была выявлена во всех группах. Но плотность и интенсивность ее была различной.
Максимальное снижение экспрессии серотонина определялось в ЧЛО I группы. Так, в центральных участках площадь экспрессии серотонина была в 6,8 раз меньше по сравнению с группой сравнения и составила 0,6±0.01% (р<0.001). В периферических отделах также было выявлено его достоверное снижение. В I группе этот показатель составил 07±0.01%. во 2 группе – 2,2±0,02% (р<0.001). Как в центральных, так и в периферических участках ЧЛО I группы экспрессия серотонина была в 2 раза ниже по сравнению со 2 группой.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
