
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Проведено эпидемиологическое исследование, которое выявило взаимосвязь микоплазменной инфекции (M. hominis) с раком простаты.
Полученные результаты показывают необходимость проведения дополнительных диагностических и профилактических мероприятий, направленных на идентификацию и лечение микоплазменных инфекций, особенно, у больных, которым показана химиотерапия.
Показано, что анти-апоптотическая активность микоплазменных диациллипопептидов может быть использована для создания радиопротективных средств, а также средств для мобилизации клеток предшественников гемопоэза, что может найти широкое применение в трансплантологии.
Отдельные положения работы включены в:
· Методические рекомендации «Определение бактериальных эндотоксинов в биопрепаратах при использовании клеточной линии 293, экспрессирующей Толл-подобный рецептор 4». М.-2009.
· Использование методов количественного определения наноматериалов на предприятиях наноиндустрии. Методические рекомендации. М.: Федеральный Центр гигиены и эпидемиологии Роспотребнадзора, 2010. МР 1.2.2639-10
· Медико-биологическая оценка безопасности наноматериалов. Методические указания М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010. МУ 1
· Токсиколого-гигиеническая оценка безопасности наноматериалов. Методические указания М.: Федеральный Центр гигиены и эпидемиологии Роспотребнадзора, 2009. МУ 1.2.2520-09
Положения выносимые на защиту
Микоплазменная инфекция вызывает изменения активности ключевых транскрипционных факторов р53 и NF-kB. Это приводит к изменению восприимчивости инфицированных клеток к различным проапоптотическим стимулам, а также способствует их необратимой Ras-зависимой трансформации.
Активация NF-kB транскрипционного фактора в опухолевых клетках в ответ на микоплазменную инфекцию (или ее структурные компоненты) является существенным фактором прогрессии опухолевого роста и снижения чувствительности опухолевых клеток к химиотерапии в условиях in vivo.
Полученные данные, характеризующие трансформирующую активность микоплазм, а также влияние их компонентов на прогрессию опухолевого роста, являются существенными для понимания механизма взаимодействия бактериальных патогенов с клетками хозяина, а также для выяснения роли хронических бактериальных инфекций в прогрессии опухолевых заболеваний.
Апробация работы
Основные результаты работы были доложены и обсуждены на XII международной конференции "СПИД, рак и родственные проблемы", 2004 г., IX конгрессе Всероссийского научно-практического общества эпидемиологов, микробиологов и паразитологов, 2007 г., Всероссийской научно-практической конференции «Вакцинология 2006», Российской научной конференции с международным участием «Фундаментальные исследования в уронефрологии» 2009 г., XIII Российском онкологическом конгрессе, 2009 г. IV Российском симпозиуме «Белки и пептиды», 2009 г.
Результаты работы были доложены и обсуждены на расширенном заседании Ученого совета ФГБУ «НИИЭМ им. » Минздравсоцразвития России, 2007 г. и на научной конфереции отдела Биологии стресса клетки института рака Розвелла Парка, Баффало, США, 2008 г.
Апробация диссертации состоялась 23.11.10 на научной конференции отделов генетики и молекулярной биологии бактерий и медицинской микробиологии ФГБУ «НИИЭМ им. » Минздравсоцразвития России
Публикации
По материалам диссертации было опубликовано 18 научных работ, в том числе 13 в журналах рекомендованных ВАК РФ.
Полученные результаты нашли отражение в 4 патентах на изобретения.
Объем и структура диссертации
Диссертационная работа изложена на 226 листах машинописного текста и включает Введение и 4 главы: «Обзор литературы», «Материалы и методы», «Результаты исследований», «Обсуждение результатов», Выводы, Список используемой литературы (265 источников, из которых 7 отечественных и 258 иностранных). Работа содержит 5 таблиц и 44 рисунка.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследований
Экспериментальное заражение культур клеток различными видами микоплазм, а также идентификация и количественная оценка микоплазм в инфицированных клетках была выполнена совместно с лабораторией микоплазм и Л-форм бактерий ФГБУ НИИЭМ им. Минздравсоцразвития РФ. Эксперименты по изучению способности микоплазм модулировать активность транскрипционных факторов NF-kB и р53 была выполнена совместно с (Roswell Park Cancer Institute, Buffalo NY).
В работе были использованы:
– штаммы микоплазм M. arginini, M. orale, M. hominis, M. fermentans, M. pneumoniae были любезно предоставлены д. б.н. (лаборатория микоплазм и Л-форм бактерий ФГБУ НИИЭМ им. Минздравсоцразвития РФ).
– плазмида pLV-CMV-Super-repressor-Bleo, содержащая мутантный IkB, была любезно предоставлена (Roswell Park Cancer Institute, Buffalo NY).
– лабораторные штаммы Escherichia coli DH5a;
- клеточные линии MCF-7, HCT-116, содержащие ген b-галактозидазы под контролем р53 зависимого промотера, а также клеточные линии 293-null, 293hTLR2, 293hTLR1/2, 293hTLR2/6, 293hTLR2/CD14, 293hTLR4/MD2-CD14 (клетки эмбриональной почки человека, содержащие ген b-галактозидазы под контролем NF-kB-зависимого промотора, и экспрессирующие Толл-подобные рецепторы TLR2, TLR2/6, TLR2/CD14, TLR4/MD2-CD14) были любезно предоставлены А.В. Гудковым (Roswell Park Cancer Institute, Buffalo NY).
– коммерческие препараты специфических эндодезоксирибонуклеаз и других ферментов фирм "Fermentas MBI" (Литва), "Promega" (США) и «NE BioLabs» (США). Для постановки реакции обратной транскрипции использовали набор Reverse Tpanscription System («Invitrogen», США). Для анализа концентраций цитокинов использовали набор FlowCytomix BenderMedsystems (Австрия). Синтез праймеров был выполнен в . Секвенирование проводилось в НПФ «Литех».
Эксперименты проводились на бестимусных мышах линии D2&I. и BALB/c.
- наращивание и культивирование микоплазм, а также заражение культур клеток различными штаммами микоплазм проводили по стандартным методам, изложенным в работах и (1968г.) Идентификацию и количественную оценку микоплазм в инфицированных культурах клеток проводили с использованием методов измерения активности ферментов микоплазмы в культуральной среде; ПЦР с праймерами на класс Mollicute и видоспецифическими праймерами; ПЦР в режиме реального времени.
- все генно-инженерные манипуляции проводились по стандартным методам, изложенным в Maniatis Т. (1982). Анализ активности транскрипционного фактора р53 и NF-kB в клетках проводили спектрофотометрически по реакции на β-галактозидазу методом ОТ-ПЦР и методом иммуноблоттинга. Анализ пролиферации, выживаемости и уровня апоптоза в клетках проводили по реакции с субстратом МТТ, окраской клеток метиленовым голубым; измерением активности каспаз 3/7, спектрофотометрически, используя специфический флюорогенный субстрат Ac-DEVD-AMC; Анализ экспрессии Толл-подобных рецепторов 2 и 6 в опухолевых клетках проводили методом обратной транскрипции с последующей ПЦР на гены Толл-подобных рецепторов 2 и 6 и методом проточной цитофлюорометрии. Анализ экспрессии цитокинов проводили с использованием набора BenderMedsystems (Австрия) при помощи проточного цитофлюориметра с использованием коммерческого набора FlowCytomix
- статистическую обработку результатов проводили общепринятыми методами с использованием программного обеспечения Microsoft Excel.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Изучение влияния различных видов микоплазм на транскрипционную активность р53
Для проверки предположения о способности микоплазм ингибировать активность р53, была использована репортерная линия клеток (Balb3T3), несущая в своем геноме p53-респонсивный элемент, под контролем которого находится ген b-галактозидазы. Такая клеточная линия позволяет производить количественный анализ активности белка р53, используя стандартную цветную реакцию с ONPG.
Данная репортерная линия заражалась Mycoplasma sp. и далее обрабатывалась доксорубицином (Dox) для индукции р53. После чего, как в обработанных Dox, так и в необработанных Dox клетках (зараженных различными видами микоплазм и контрольных незараженных клетках) оценивалась активность р53 по изменению уровня активности b-галактозидазы. Измерение активности b-галактозидазы производили через 24 часа после добавления Dox (рис.1).
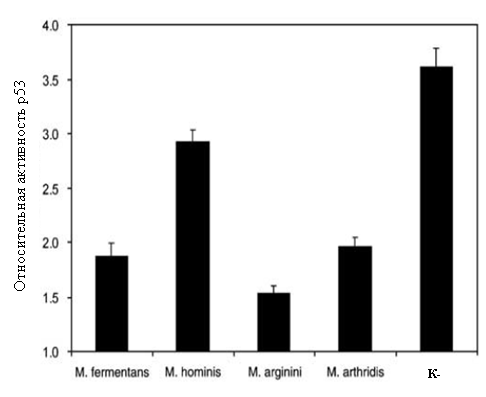 |
Рисунок 1. Влияние микоплазменной инфекции на активность транскрипционного
фактора р53.
Относительная активность определялась по реакции ONPG как отношение значения оптической плотности в экспериментальном образце,(клетках обработанных доксорубицином, к значению оптической плотности в контрольном образце (клетках необработанных доксорубицином). Доксорубицин в культуральную среду добавляли до концентрации 1мг/мл. Клетки с доксорубицином культивировали в течение 24 часов. р53-зависимую индукцию b-галактозидазы определяли спектрофотометрически (414 нм) по изменению оптической плотности красящего раствора, содержащего о-нитрофенил-b-D-галактопиранозид (ONPG).
Полученные данные показали, что все использованные виды микоплазм способны супрессировать активность р53. Наибольшее ингибирование р53 наблюдалось в клетках, инфицированных микоплазмой M.arginini. Этот вид микоплазмы был выбран в качестве модели для проведения дальнейших исследований.
На следующем этапе M.arginini была использована для более детального исследования влияния микоплазменной инфекции на транскрипционную активность р53. Для дальнейшего более детального изучения влияния микоплазменной инфекции на транскрипционную активность р53, M.arginini были инфицированы человеческие клеточные линии HCT-116/ConA и MCF-7/ConA, также несущие р53-респонсивный элемент с геном b-галактозидазы [Gurova К., 2005]. Активность р53 была оценена в сравнении с неинфицированными клетками по изменению уровня активности репортера в ответ на обработку клеток ДНК-повреждающим агентом 5-фторурацилом (5-ФУ). Как видно из полученных результатов (Рис. 2А.) в инфицированных клетках наблюдалось очевидное снижение активности р53, при том, что выживаемость инфицированных и неинфицированных клеток статистически не отличалась.
В дополнение к трансактивационному анализу, подавление активности р53 в условиях микоплазменной инфекции, было охарактеризовано по изменению уровня транскрипции эндогенного р53-регулируемого гена – p21waf1. Детекция мРНК этого гена проводилась с помощью РТ-ПЦР. В результате этого эксперимента было показано снижение уровня индукции мРНК гена р21 в ответ на повреждение ДНК, вызванное 5-ФУ, в клетках инфицированных M.аrginini (Рис 2Б). Далее для характеристики р53-ингибирующей активности M.arginini былo проанализированo изменение уровня экспрессии p53-респонсивного гена p21 в клетках с эндогенным (диплоидные фибробласты человека BJ, обработанные 5-ФУ) и эктопически экспрессированным р53 (р53-негативные клетки аденокарциномы легкого человека H1299, инфицированные рекомбинантным аденовирусом человека 5 серотипа, экспрессирующим р53).
Как видно из Рис 2В. микоплазменная инфекция вызывала существенное снижение уровня индукции белка р21 в обеих клеточных системах. Работа проводилась совместно с к. б.н.
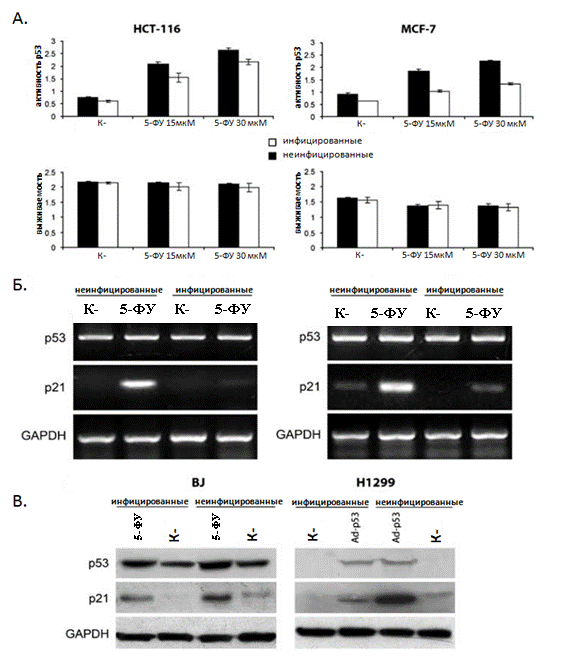
Рисунок 2. Подавление транскрипционной активности р53 микоплазмой M.arginini
(А) Активность р53-респонсивного репортера в контрольных и инфицированных M. arginini клетках HCT-116 и MCF-7. HCT-116 и MCF-7 клетки, стабильно трансфицированные лентивирусным вектором, несущим р53-респонсивную конструкцию ConALacZ , были посеяны на 96-луночный планшет, после чего инкубировались 16 часов в среде, содержащей указанные на рисунке концентрации 5-ФУ. Клетки инфицировали M.arginini не менее чем за 2 недели до эксперимента. Присутствие M. arginini в клеточной культуре подтверждали высевом, ПЦР и набором MycoAlert. Активность репортера (b-галактозидазы) определяли по конверсии ONPG (верхняя панель графиков), выживаемость клеток оценивали по окраске метиленовым голубым (нижняя панель графиков).
(Б) РТ-ПЦР анализ экспрессии p53 и p21 в контрольных и инфицированных M. arginini клетках HCT-116 и MCF-7. Для индукции р53 клетки обрабатывали 5-ФУ (30 мкM) в течение 24 часов. Уровень экспрессии мРНК p53 и p21 оценивали методом РТ-ПЦР. Уровень экспрессии GAPDH (glyceraldehyde-3-phosphate dehydrogenase) использовался как положительный контроль, подтверждающий схожее качество и количество кДНК в каждой пробе. Клетки HCT-116 - левая электрофореграмма, MCF-7 правая электрофореграмма .
(В) Влияние микоплазменной инфекции (M. arginini) на уровень экспрессии р53-респонсивного гена р21. Лизаты, приготовленные из контрольных (неинфицированных) и инфицированных M. arginini клеток BJ и H1299 были использованы для Вестерн-блот анализа уровня экспрессии белков p53 и p21. Уровень экспрессим GAPDH использовался как контроль внесения общего количества белка в каждый трек. Для индукции р53 клетки BJ брабатывали 5-ФУ (30 мкМ) в течение 16 часов, а клетки H1299 обрабатывали аденовирусом, экспрессирующим p53 в количестве 10 БОЕ (бляшкообразующих единиц) на клетку.
Микоплазменная инфекция ингибирует р53-зависимую остановку клеточного цикла и р53-зависимый апоптоз
р53 является одним из ключевых белков, определяющих прохождение клетки через сверочные точки клеточного цикла. Для проверки влияния микоплазменной инфекции на эту функцию р53 был поставлен эксперимент по анализу включения бромдезоксиуридина (BrdU) в ДНК р53-позитивных клеток линии REF52, инфицированных и неинфицированных M.arginini. Клетки этой линии представляют собой иммортализованные фибробласты эмбриона крысы, которые способны подвергаться р53-зависимому аресту G1/S сверочной точке клеточного цикла [Sablina А., 1999]. Культура клеток REF52 была синхронизирована в бессывороточной среде в течение 72 часов, после чего обрабатывалась ДНК-повреждающим агентом – 5-ФУ.
Как можно увидеть на Рис. 3А, только 2% неинфицированных клеток смогли преодолеть запрет на вхождение в фазу синтеза (1.2% из всех клеток, обработанных 5-ФУ были BrdU-позитивными, при этом в контрольном варианте необработанном 5-ФУ процент BrdU-позитивных клеток составил приблизительно 60%). При этом в популяции инфицированных клеток около 13% клеток преодолели сверочную точку и вошли в S-фазу (8% из всех клеток, обработанных 5-ФУ были BrdU-позитивными, при этом в контрольном варианте необработанном 5-ФУ процент BrdU-позитивных клеток составил приблизительно 60%). Таким образом, полученные результаты показывают, что микоплазменнная инфекция способна ослаблять р53-зависимый арест, индуцированный обработкой 5-ФУ. Другой важной функцией, которую выполняет р53, является индукция апоптоза. Для оценки способности микоплазменной инфекции ингибировать р53-зависимый апоптоз в эксперименте была использована р53-негативная клеточная линия H1299, которая характеризуется быстрой индукцией апоптоза в ответ на восстановление функции р53. Клетки линии Н1299, инфицированные и неинфицированные M. arginini были трансдуцированы рекомбинантным аденовирусом человека, экспрессирующим дикий тип гена р53 (Ad-p53). Для контроля эффективности трансдукции в эксперименте использовали рекомбинантный аденовирус, экспрессирующий репортерный ген SEAP (placental-secreted alkaline phosphatase). Из данных представленных на Рис. 15Б очевидно, что активность SEAP была схожей в инфицированных и неинфицированных микоплазмой клетках. Это доказывает, что микоплазменная
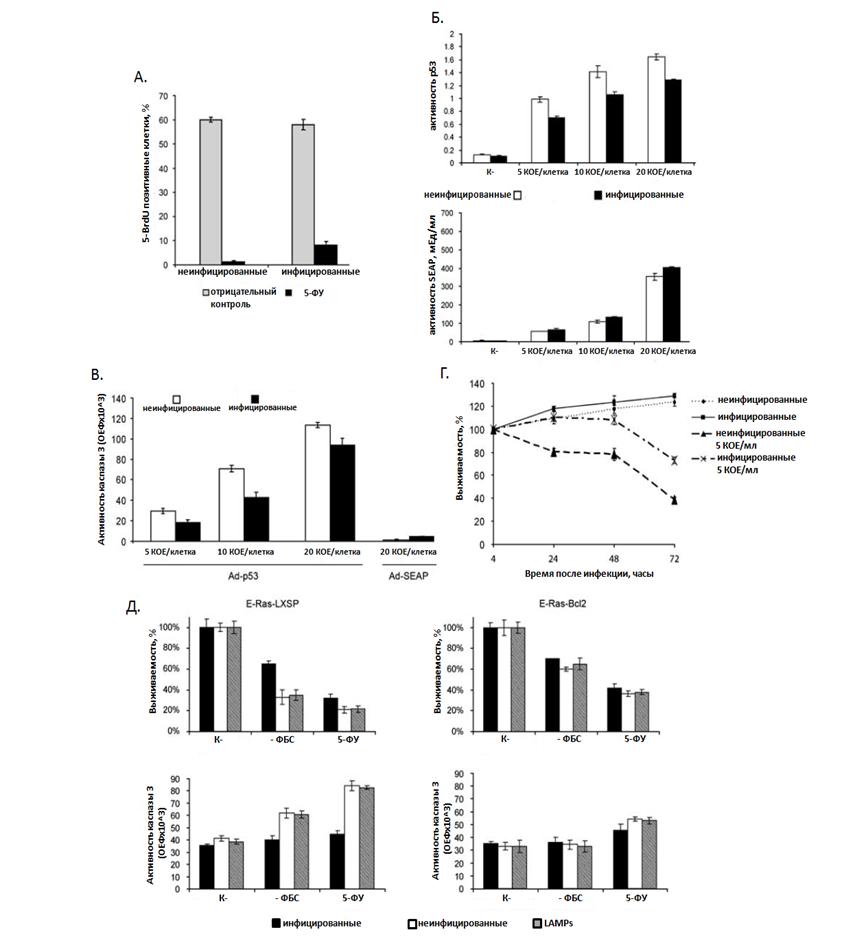
Рисунок 3. Инфекция M. arginini ингибирует р53-зависимый арест клеточного цикла и р53-зависимый апоптоз.
(А.) Эффект микоплазменной инфекции на способность клеток REF52, синхронизированных в G0/G1 фазе на бессывороточной среде (в течение 72 часов) входить в S-фазу (включать 5-бромдезоксиуридин (5-BrdU) в растущую цепь ДНК). Включение 5-BrdU оценивали методом иммунофлюоресценции, используя специфические антитела к BrdU. Оценка включения BrdU проводилась через 7 часов после стимуляции клеток средой, содержащей 10% фетальной бычей сыворотки (ФБС) и 5-BrdU (отрицательный контроль) или 5-BrdU с 5-ФУ. Для подсчета в каждом варианте учитывали данные не менее чем для 500 клеток.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
