
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
(Б.) Средняя масса селезенки. Селезенки извлекали из анестезированных мышей. Экспериментальные группы мышей идентичны описанным выше. Для определения средней массы использовали по 5 органов из каждой группы мышей.
(В.) Микрофотографии срезов печени. Извлеченные образцы печени помещали в 10% формалин для фиксации. Далее по стандартной методике образцы заключали в парафин, приготовленные срезы окрашивали гематоксилин-эозином.
(Г.) Диаграмма выживаемости мышей линии Balb/С . WEHI-3В K- группа мышей с внутрибрюшинным введением клеток WEHI-3B; WEHI-3В + R-Pam2 - группа мышей с внутрибрюшинным введением клеток WEHI-3B и внутримышечным введением R-Pam2; WEHI-3В + 5-Fu - группа мышей с внутрибрюшинным введением клеток WEHI-3B, получающие химиотерапевтический препарат 5-фторурацил; WEHI-3В + R-Pam2 + 5-Fu - группа мышей с внутрибрюшинным введением клеток WEHI-3B и внутримышечным введением R-Pam2, получающие химиотерапевтический препарат 5-фторурацил. (р<0,001 по лог-ранговому тесту).
Работа выполнена совместно с к. б.н. [лаборатория молекулярной биотехнологии ФГБУ «НИИЭМ им. » Минздравсоцразвития России]
Изучение влияния структурного компонента M. arginini (R-Pam2) на продукцию факторов, стимулирующих пролиферацию клеток мышиной миеломоноцитарной лейкемии WEHI-3B in vivo
Принимая во внимание основное отличие результатов экспериментов in vitro и in vivo по скорости роста клеток WEHI-3B в присутствии R-Pam2, мы предположили, что в организме экспериментальных животных после введения диациллипопептида M.arginini может происходить выработка факторов, которые являются существенными для пролиферации клеток WEHI-3B, тем самым определяя ускоренную кинетику их роста in vivo.
Для подтверждения выдвинутой гипотезы в эксперименте in vitro мы изучили влияние добавления к клеткам WEHI-3B сыворотки мышей, получавших R-Pam2 на скорость их пролиферации. Для этого мышам линии BALB/c внутримышечно вводили диациллипопептид M.arginini R-Pam2 в концентрации 5 мкг/мышь. Через 24 часа после инъекции у мышей проводили отбор крови для получения сыворотки. В качестве контроля использовали сыворотку мышей, получавших солевой фосфатный буфер. Полученные сыворотки использовали для приготовления 5% культуральной среды для клеток WEHI-3B (RPMI). Среды добавляли к клеткам WEHI-3B и высаживали на 96-луночный планшет в концентрации 103 клеток/лунку. Кинетику роста клеток определяли по накоплению клеточной биомассы в реакции с субстратом МТТ в течение 72 часов (рисунок 12).
Как видно из рисунка 12, добавление к клеткам WEHI-3B сыворотки крови мышей, получавших внутримышечно R-Pam2, способствовало увеличению скорости их пролиферации. Результаты, полученные в данном эксперименте, подтвердили сделанное ранее предположение о возможной выработке фактора, индуцирующего рост клеток мышиной миеломоноцитарной лейкемии в ответ на введение R-Pam2.

Рисунок 12. Кинетика роста клеток мышиной миеломоноцитарной лейкемии WEHI-3B при добавлении сыворотки мышей, получавших R-Pam2 (р<0,005)
На следующем этапе была предпринята попытка идентификации кандидатных факторов, способствующих усилению роста клеток WEHI-3B.
Для выполнения поставленной задачи нам был проанализирован спектр экспрессии хемокинов и цитокинов в организме животного в ответ на введение диациллипопептида M.arginini (рис.12А.). Мышам линии BALB/c внутримышечно вводили R-Pam2 в количестве 5 мкг/мышь. Далее, у мышей отбирали кровь и в полученной сыворотке методом проточной цитофлюориметрии с использованием набора FlowCytomix BenderMedsystems (Австрия) определяли концентрации 15 хемокинов и цитокинов (IL-1,2,4,5,6,10,17, ФНОа, ГМ-КСФ, MCP-1,3 MIP-1a,1b, RANTES, ИФН-гамма). На рисунке 13 представлены данные для цитокинов, уровень экспрессии которых изменялся в ответ на введение R-Pam2. Как видно из рисунка, в ответ на введение R-Pam2, изменялась экспрессия 8 из 15 исследованных цитокинов. Анализ литературных данных показал, что 5 из этих цитокинов– IL-6, MCP-1, MCP-3, RANTES, ФНОа, способны напрямую или опосредованно стимулировать опухолевый рост. Следует отметить, что полученные данные были ограничены только 15 измеряемыми цитокинами, тогда как на рост опухоли в организме может влиять значительно большее количество факторов. В связи с этим, для выявления дополнительных опухолестимулирующих факторов мы провели исследование in vivo в котором были определены органы, вовлеченные в ответ на системное циркулирование диациллипопептидов, а также проведен микроэррей-анализ для определения изменений в профиле экспрессии генов в ответ на действие микоплазменных антигенов. На первом этапе были использованы
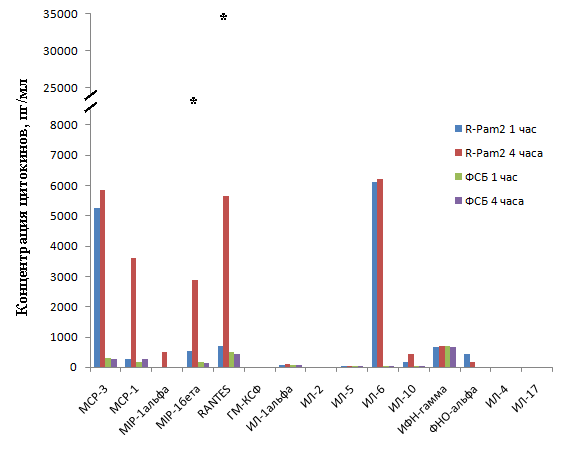 |
Рисунок 13. Определение концентрации цитокинов в сыворотках крови мышей, инъецированных R-Pam2. Мышам линии BALB/C вводили диациллипопептид R-Pam2 или фосфатно-солевой буфер (ФСБ), после чего через 0,33; 0,66; 1, 2, 4, 6, 16, 24, 72 часа отбирали кровь и приготавливали сыворотку. Полученные образцы сыворотки использовали для определения уровня экспрессии цитокинов. Каждая точка представляет собой среднее значение из трех независимых повторов. На графике представлены данные соответствующие пиковой концентрации цитокинов – 1 или 4 часа после введения R-Pam2. *- Для MIP-1альфа и RANTES представлены значения концентраций соответствующие 4 и 6 часам после инъекции R-Pam2 - точкам появления и пика концентраций этих цитокинов в сыворотке крови
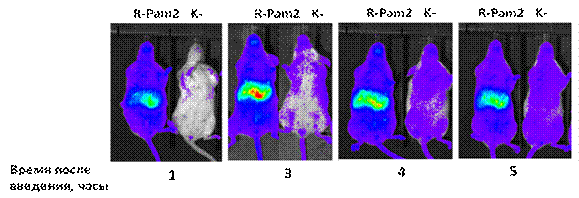
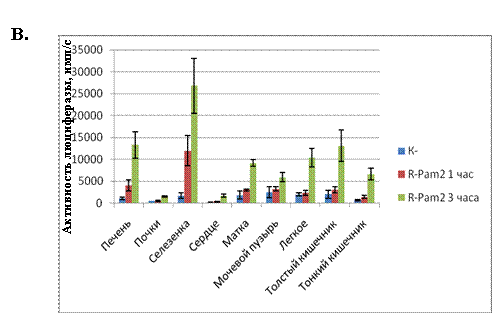
трансгенные мыши (полученные на основе линии Balb/C), содержащие в своем геноме NF-kB-зависимый промотер (промотер ген IkB) под контролем которого находится ген люциферазы. Мышам вводили синтетический диациллипопептид R-Pam2 (5мкг/мышь), после чего через указанные на рисунке 14А. интервалы времени, регистрировали биолюминисценцию (NF-kB-зависимую экспрессию люциферазы). Как видно из результатов в организме мышей развивался очень быстрый ответ, с максимумом активации NF-kB через 3 часа после введения R-Pam2. Для определения активности NF-kB в отдельных органах, трансгенным мышам вводили R-Pam2 после чего, через 1 и 3 часа, отбирали органы указанные на рисунке 14Б. Из отобранных органов приготавливали экстракт, после чего в экстрактах нормализованных по концентрации белка (30 мкг белка на реакцию), определяли уровень активности NF-kB. Полученные данные показали, что введение R-Pam2 вызывает достоверную активацию NF-kB во всех отобранных органах (см. рис.14В.). То есть, из полученных результатов можно сделать заключение, что диациллипопептиды способны оказывать системное действие на организм. В дополнительных экспериментах (данные не приводятся) было показано, что активация NF-kB также наблюдается в поджелудочной железе, лимфоузлах, незначительно, в костном мозге и мышцах. Практически не детектируемый уровень активации NF-kB наблюдался в образцах мозга и кожи. Следует отметить, что наибольшая активация NF-kB была обнаружена в двух органах - селезенке и печени. Наблюдаемая активность NF-kB в печени и селезенке хорошо согласуется с данными по метастазированию клеток WEHI-3B именно в эти органы.
На следующем этапе исследования для выявления дополнительных факторов, способных влиять на прогрессию опухоли мы определили, каким образом меняется спектр экспрессии генов в ответ на введение R-Pam2. Для этого была использована линия мышей Balb/C. Мышам вводили R-Pam2 (5 мкг/мышь) после чего через час отбирали образцы печени. Точка отбора образцов (1 час) для определения профиля экспрессии мРНК была выбрана с учетом того, что максимум NF-kB-зависимой экспрессии люциферазы (белка) приходится на 3 часа после введения R-Pam2. Выбор печени в качестве исследуемого органа был обусловлен тем, что она является (наряду с селезенкой) наиболее респонсивным в отношении P-Pam2 органом. Кроме того, печень является еще и самым крупным органом, а следовательно, ее активация может иметь наибольшее значение для организма в целом.
Отобранные образцы печени передавали в специализированной подразделение Института рака Розвелла Парка, где с использованием микрочипов фирмы Illumina, Inc (США) производилось полногеномное исследование изменения экспрессии генов. Проведенный анализ полученных результатов показал, что введение в организм диациллипопептида микоплазмы R-Pam2 повышает более чем 2 раза экспрессию свыше 3000 генов, экспрессия более чем 500 из этих генов
повысилась в 5-400 раз.
Ниже в таблице 2 приведен перечень генов, экспрессия которых способна позитивно влиять на рост опухоли. Кроме того, в таблицу внесены гены, участвующие в развитии провоспалительных и антиапототических реакций, которые также опосредовано могут быть важны для опухолевой прогрессии.
А.
 | |
| |

Рисунок 14. Влияние диациллипопептидов микоплазм на активность транскрипционного фактора NF-kB в различных органах мышей.
(А.) Изменение активности NF-кВ в трансгенных мышах в зависимости от времени.
Трансгенным мышам подкожно вводили 5 мкг R-Pam2 или в качестве контроля фосфатно-солевой буфер (ФСБ). Далее, непосредственно перед измерением уровня NF-kB-зависимой экспрессии люциферазы, мышам внутрибрюшинно вводили 20 мкг люциферина. Через 5 минут после введения опытных и контрольных мышей анестезировали изофлюраном и помещали в специальный детектор биолюминисценции (Xenogene, USA), где через указанные на рисунке интервалы времени определяли активность люциферазы
(Б.) Схема отбора органов из трансгенных мышей.
Трансгенных мышей анестезировали изофлюраном, производили цервикальную дислокацию и отбирали указанные на рисунке органы в пробирки, помещенные в сухой лед.
(В.) NF-kB-зависимая экспрессия люциферазы в органах трансгенных мышей, инъецированных диациллипопептидами микоплазмы.
Помещенные в сухой лед органы, переносили в лед, добавляли ФСБ, содержащий коктейль протеазных ингибиторов и гомогенизировали. Гомогенаты (супернатанты) использовали для определения активности люциферазы.
Данные микроэррей-анализа, объединенные с данными определения спектра цитокинов, показывают, что введение R-Pam2, приводит к синтезу в организме целого ряда белков, для которых показана опухолестимулирующая
активность: MCP-1, MCP-3, Gro1, RANTES, IL-1, IL-6, ФНОа, S100A9, SAA1, SAA3 и др. По-видимому, именно совокупное действие перечисленных факторов способствует более быстрому росту опухолей WEHI-3B in vivo.
Таблица 2. Изменение экспрессии провоспалительных и антиапоптотических белков в клетках печени в ответ на введение R-Pam2.
Краткое название гена | Полное название гена | Изменение уровня экспрессии, разы |
Цитокины: CXCL1 (GRO1) CXCL10 (IP10) CXCL2 (GRO2 CXCL9 (MIG) CCL4 (MIP-1-beta) CCL7 (MCP-3) IL1A IL1B TNFа | хемокин, содержащий (C-X-C мотив) 1 хемокин, содержащий (C-X-C мотив) 10 хемокин, содержащий (C-X-C мотив) 2) хемокин, содержащий (C-X-C мотив) 9 хемокин, содержащий (C-C мотив) 4 хемокин, содержащий (C-C мотив) 7 интерлейкин 1 альфа интерлейкин 1 бета Фактор некроза опухоли альфа | 39.8 36.1 54.4 3.4 226.7 118.2 23.4 62.1 174.0 |
Паттерн-распознающие и их адаптерные молекулы: TLR2 FCNA NOD1 SAA1 SAA3 CD14 MYD88 | Толл-подобный рецептор 2 Фиколин Нуклеотид-связывающий и содержащий домен олигомеризации белок Сывороточный амилоид А1 Сывороточный амилоид А3 Молекула CD14 Ген первичного ответа миелоидной дифференцировки(88) | 4,3 2,9 2.0 3.1 5.5 41.4 192.9 |
Антимикробные белки CFP HC LCN2 S100A8 S100A9 | Фактор комплемента пропердин Гемолитический комплемент Липокалин 2 S100 кальций-связывающий белок А8 S100 кальций-связывающий белок А9 | 3.0 2.9 444.6 65.6 14.8 |
Антиапоптотические белки IER3 MCL1 Bcl2 BclXl BCL2A1B BCL2A1D SUPV3L1 | Белок немедленно раннего ответа 3 Миелоидных лейкозных клеток последовательность 1 B-клеточная лейкемия/лимфома 2 (Bcl2) Bcl2-родственный белок Xl Bcl2-родственный белок А1В Bcl2-родственный белок А1D Супрессор var1, 3-подобный 1 | 17.7 1.9 1,5 1,7 14.7 3.3 2.9 |
Возможно, что опухолестимулирующим действием могут обладать и другие белки, продукция которых меняется в ответ на введение R-Pam2, однако в доступной научной литературе мы не обнаружили данных, подтверждающих участие этих белков в опухолевой прогрессии. Таким образом, обобщив полученные результаты, мы можем сделать вывод, что введение R-Pam2 в организм может оказывать прямое и опосредованное действие на клетки опухоли. Опосредованное действие связано с непрямым влиянием R-Pam2 на клетки опухоли, которое реализуется через TLR2-зависимую индукцию факторами, изменяющих пролиферативный потенциал опухолевых клеток и влияющих на их инвазивные свойства. В свою очередь, прямой эффект R-Pam2 связан с TLR2-зависимой активацией NF-kB и увеличением резистентности опухолевых клеток к химиотерапевтическим препаратам. При этом индукция NF-kB в клетках опухоли не измененяет их митогенную активность. Кроме того, мы показали, что сами клетки WEHI-3B в ответ на обработку R-Pam2 увеличивают экспрессию двух цитокинов - ФНОа и ИЛ-6, которые способны стимулировать опухолевую прогрессию (см. приложение 2).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
