
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Результаты проведенного исследования, как и ожидалось, показали, что микоплазменная инфекция снижает экспрессию ряда р53-зависимых генов. Особенно, примечательным выглядит снижение экспрессии гена Fas, экспрессия которого является важным фактором гибели клеток, обработанных 5-ФУ. Проведенный анализ выявил значительное количество р53-зависимых генов (tumor protein p53 inducible nuclear protein 1 (TP53INP1), BAX, TP53 regulated inhibitor of apoptosis 1 (TRIAP1), sestrin 1, sestrin2, damage-specific DNA binding protein 2 (DDB2), stratifin (SFN) и др.) экспрессия которых, в клетках инфицированных микоплазмой, подавлялась слабо (1,07-1,2 раза).
Следует отметить, что в клетках инфицированных микоплазмой, значительно (более чем в 2 раза) изменялась р53-независимая экспрессия генов (экспрессия более чем 240 генов была повышена, экспрессия более 190 генов понижена). Одним из наиболее интересных р53-независимых генов, экспрессия которого была повышена в инфицированных клетках HCT-116, является ген белка versican. В недавней работе [Kim Н., 2009] показано, что этот белок является одним из ключевых факторов метастазирования клеток рака легкого.
Различные виды микоплазм приводят к активации NF-kB в клетках при взаимодействии с Толл-подобным рецептором 2.
Целью второй части работы было изучение влияния микоплазменной инфекции на активность другого стресс-активируемого транскрипционного фактора NF-kB. Этот фактор, в отличие от р53, контролирует экспрессию анти-апоптотических белков и является одним из основных факторов прогрессии опухолевого роста. Для характеристики целого ряда эффектов, сопровождающих микоплазменную инфекцию (генетическая нестабильность, увеличение резистентности к апоптозу, стимуляция метастазирования и др.) мы решили изучить, каким образом изменяется активность NF-kB в инфицированных микоплазмой клетках.
Известно, что белок M.fermentans MALP-2 активирует NF-kB через TLR2/6, липид-ассоциированные мембранные белки (ЛАМБ) M.penetrans через TLR1/2, ЛАМБ M.pneumoniae через оба сочетания Толл-подобных рецепторов (TLR2/6 и TLR1/2), а суперантиген M.arthritidis через TLR2 и TLR4- зависимые пути [Shimizu 2004, 2005, Mu 2006]. Как видно из представленных работ наиболее универсальным набором Толл-подобных рецепторов, участвующих в распознавании структурных компонентов микоплазм, является сочетание TLR2/6. Поэтому, для изучения способности различных видов микоплазм или их структурных компонентов активировать транскрипционный фактор NF-kB была использована клеточная линия, экспрессирующая Толл-подобные рецепторы 2/6, полученная на основе клеток эмбрионального почечного эпителия человека линии 293. Дополнительно, для визуального контроля активации NF-kB методом лентивирусной трансфекции в указанные клетки был введен ген b-галактозидазы под контролем NF-kB-зависимого промотора. Клеточная линия были предоставлены (Roswell Park Cancer Institute, Buffalo NY).
Принцип использования клеточной линии основан на способности Толл-подобных рецепторов, после связывания со специфическими лигандами M.arginini, рекрутировать адаптерные молекулы MyD88, TRAF6, активировать каскад киназ, в результате чего происходит активация транскрипционного фактора NF-kB, его транслокация в ядро, где этот фактор связывается с собственным респонсивным элементом, под транскрипционным контролем которого находится ген галактозидазы. Таким образом, используемая система позволяет по цветной реакции на галактозидазу измерять активность NF-kB в клетках и регистрировать взаимодействие Толл-подобных рецепторов с лигандами микоплазмы. Указанные клетки обрабатывали либо липид-ассоциированными мембранными белками, либо культурами микоплазм (M.arginini, M.hominis, M.orale, M.fermentans, M.pneumonia). После чего проводили измерение активности NF-kB в клетках. В качестве положительного контроля активации NF-kB использовали ФНО-a (фактор некроза опухоли) (рисунок 5).
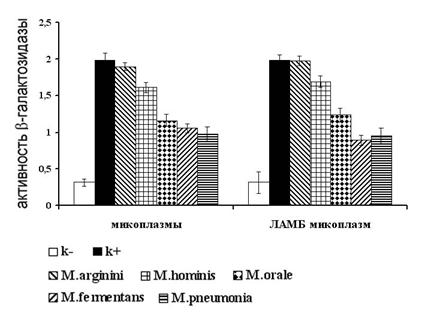
Рисунок 5. Измерение активности транскрипционного фактора NF-kB в клетках, экспрессирующих Толл-подобные рецепторы 2 и 6. Данные по каждой точке являются результатом трех независимых экспериментов. (р<0,005)
Как видно из рисунка 5 обработка клеток как живыми микоплазмами, так и их липид-ассоциированными мембранными белками, приводила к достоверному (р<0,005) увеличению уровня NF-kB-зависимой экспрессии b-галактозидазы, что свидетельствует о способности всех, используемых в работе, микоплазм и их структурных компонентов активировать NF-kB в клетках в результате взаимодействия с TLR2/6.
Определение возможных сочетаний Толл-подобных рецепторов, вызывающих активацию транскрипционного фактора NF-kB после взаимодействия M. arginini или ее структурными компонентами
В качестве объекта исследования была выбрана M.arginini, для которой нами была показана способность подавлять активность р53. Кроме того, до настоящего времени в доступной научной литературе отсутствуют данные относительно взаимодействия M.arginini с Толл-подобными рецепторами и активации транскрипционного фактора NF-kB. В нашей работе мы решили охарактеризовать данный вид микоплазмы по NF-kB активирующей способности и определить все сочетания Толл-подобных рецепторов, являющихся для нее специфическими.
Для этого был использован набор индивидуальных клеточных сублиний, экспрессирующих одно из следующих сочетаний Толл-подобных рецепторов и акцессорных молекул MD2 и CD14: hTLR4/MD2-CD14, hTLR2, hTLR2/hTLR6, hTLR1/hTLR2, hTLR2/CD14, полученных на основе клеток эмбрионального почечного эпителия человека линии 293. Во все клетки, как и в предыдущем эксперименте для визуального контроля активации NF-kB методом лентивирусной трансфекции был введен ген b-галактозидазы под контролем NF-kB-зависимого промотора. Клеточные линии были предоставлены (Roswell Park Cancer Institute, Buffalo NY).
Описанные клеточные линии обрабатывали либо липид-ассоциированными мембранными белками M.arginini (ЛАМБ), либо культурой микоплазмы. В качестве положительного контроля активации NF-kB использовали ФНО-a (фактор некроза опухоли) (рисунок 6). Указанный цитокин после взаимодействия с собственным рецептором стимулирует транслокацию NF-kB в ядро и активирует экспрессию провоспалительных факторов [Pahl Н., 1999].
Для клеток 293hTLR4/MD2-CD14, с целью определения функциональной активности экспрессируемого ими Толл-подобного рецептора 4 в качестве дополнительного положительного контроля использовали специфический для данного рецептора лиганд - липополисахарид (ЛПС) E.coli.
Как видно из рисунка 6, обработка клеток ФНО-a приводила к достоверному (р<0,005) увеличению уровня NF-kB-зависимой экспрессии b-галактозидазы во всех линиях клеток, что подтверждает функциональную активность NF-kB-респонсивного элемента, введенного в их геном.
Обработка клеток структурными элементами микоплазмы (ЛАМБ) или самой микоплазмой, вызывала активацию NF-kB-зависимой экспрессии гена галактозидазы только в клетках, экспрессирующих следующие сочетания Толл-подобных рецепторов: hTLR2/hTLR6, hTLR1/hTLR2, hTLR2/CD14. В клетках, неэкспрессирующих Толл-подобные рецепторы или несущих hTLR4/MD2-CD14 или hTLR2, достоверного увеличения уровня NF-kB-зависмой экспрессии b-галактозидазы не детектировалось.
Примечательно, что повышенная активность транскрипционного фактора NF-kB в клетках 293hTLR2/6, 293hTLR2/CD14 и 293hTLR1/2, инфицированных M.arginini, сохранялась в течение длительного периода и наблюдалась через 1, 7 и 35 дней после инфицирования.
Эти результаты дают основание полагать, что активация данного транскрипционного фактора в клетках в ответ на микоплазменную инфекцию является конститутивной (рисунок 7).
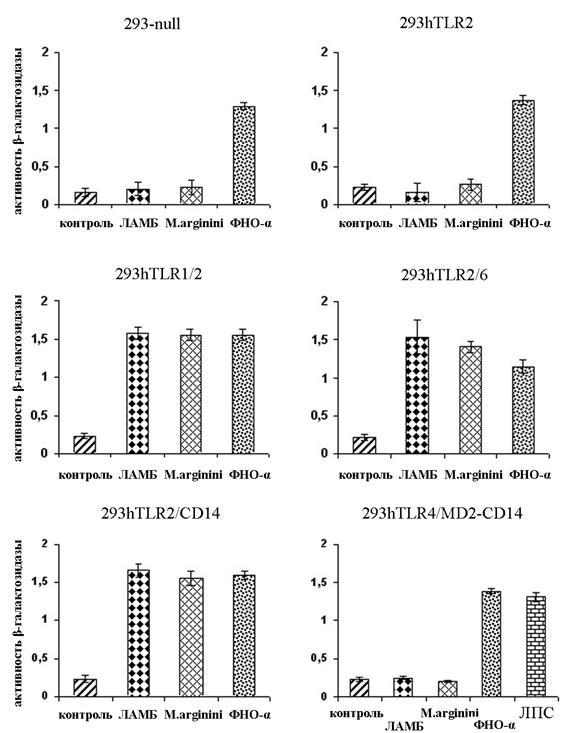
Рисунок 6. Измерение активности транскрипционного фактора NF-kB в клетках, экспрессирующих различные сочетания Толл-рецепторов. Данные по каждой точке являются результатом трех независимых экспериментов. (р<0,005)
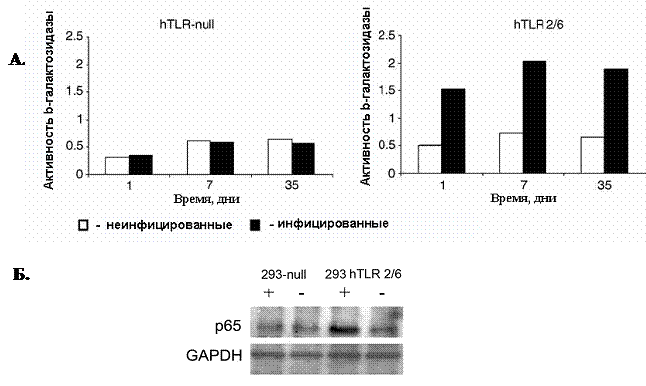
Рисунок 7. Конститутивная активация NF-kB в клетках, инфицированных микоплазмой.
(А.) Измерение активности транскрипционного фактора NF-kB в клетках в различные периоды пассирования. Данные по каждой точке являются результатом трех независимых экспериментов. (р<0,005)
(Б.) Измерение уровня активности транскрипционного фактора NF-kB в клетках, экспрессирующих Толл-подобные рецепторы TLR2/6 методом Вестерн-блот анализа. (+ клетки, инфицированные M.arginini; - клетки, неинфицированные M.arginini).
Дополнительно, активацию транскрипционного фактора NF-kB в клетках в различные периоды пассирования определяли по миграции субъединицы NF-kB, белка р65, в ядро, используя метод Вестерн-блот анализа. В клетках 293hTLR2/6, инфицированных M.arginini происходила транслокация белка р65 в ядро, в то время как в клетках, не экспрессирующих Толл-подобные рецепторы, или клетках 293hTLR2/6, неинфицированных микоплазмой, уровень белка р65 в ядре не изменялся (Рисунок 7Б.).
Таким образом, на данном этапе исследования нами было показано, что различные виды микоплазм и их липид-ассоциированные мембранные белки способны активировать транскрипционный фактор NF-kB при взаимодействии с Толл-подобными рецепторами. Дополнительно, для M.arginini были определены все сочетания Толл-рецепторов, участвующие в распознавании ее структурных компонентов и активации NF-kB. Было показано, что рецепторы TLR2/6, TLR2/CD14 и TLR1/2 одинаково эффективно распознают как клетки M.arginini, так и экстрагированные из ее мембран липопротеиды (ЛАМБ).
Работа была выполнена совместно с д. б.н. (Roswell Park Cancer Institute, Buffalo NY).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
