
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
M. arginini и ее структурные компоненты подавляют апоптоз в опухолевых клетках, экспрессирующих Толл-подобные рецепторы 2/6
На следующем этапе исследования мы решили изучить, влияет ли активация NF-kB, вызванная микоплазмами или их структурными компонентами, на выживаемость опухолевых клеток в ответ на действие химиотерапевтических препаратов.
Для этого на начальном этапе был проведен скрининг опухолевых клеточных линий (WEHI-3B, B10M, L929, U937, A549, H460, H1299, HCT-116, MCF-7) на предмет выявления экспрессии Толл-подобных рецепторов, специфичных к микоплазме. Было обнаружено, что 5 из 10 линий экспрессировали TLR2 и 6 (см. главу 3.3.5 диссертации). Для дальнейшего исследования эффектов микоплазменной инфекции и ее компонентов, в экспериментах in vitro и in vivo, нами в качестве модели была выбрана линия мышиной миеломоноцитарной лейкемии WEHI-3B. При использовании этой линии клеток было показано, что микоплазменная инфекция как и добавление R-Pam2 к опухолевым клеткам WEHI-3B, вела к активации в них транскрипционного фактора NF-kB (см. рисунок 8).
Для оценки влияния микоплазмы на апоптоз, индуцируемый химиотерапевтическими препаратами, в опухолевых клетках WEHI-3B, экспрессирующих Толл-подобные рецепторы 2 и 6, проводили измерение следующих параметров: выживаемость, уровень каспаз 3/7 (рисунок 25). Клетки инфицировали M.arginini, после чего обрабатывали циспластином, таксолом или фторурацилом. После инкубации в течении 16-18 часов проводили измерение указанных параметров.
Б. А.

|
|
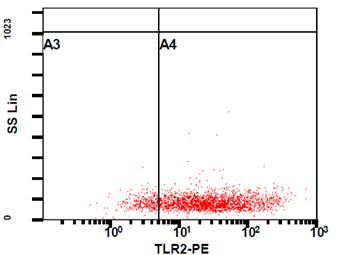
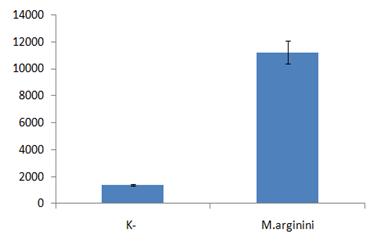
Рисунок 8. Активация NF-kB в клетках WEHI-3B в ответ на микоплазменную инфекцию.
(А.) Экспрессия TLR2 в клетках WEHI-3B. Экспрессия основного рецептора для диацилипопептидов микоплазмы была подтверждена методом проточной цитофлюорометрии. Клетки WEHI-3B инкубировали с антителами специфичными к TLR2 (eBioscience, USA). Контрольные клетки инкубировали с изотипическим контролем (анти-IgG). Исходя из фоновой флюоресценции отрицательного контроля был определен процент TLR2-позитивных клеток (измерение TLR2 проведено в полном соответствии с рекомендациями производителя).
(Б.) NF-kB-зависимая экспрессия люциферазы. В клетки WEHI-3B методом лентивирусной трансфекции был введен NF-kB-респонсивный элемент, под контролем которого находится ген люциферазы. Полученные клетки инфицировали M.arginini, после чего в инфицированных и контрольных (неинфицированных) клетках, используя стандартную методику, определяли уровень NF-kB-зависимой экспрессии люциферазы.
На рисунке 9 показана выживаемость и уровень активности основных эффекторных каспаз-3/7, при индукции апоптоза в клетках WEHI-3B, инфицированных M.arginini. Уровень каспаз-3/7 измеряли спектрофотометрически, используя специфический флюорогенный субстрат Ac-DEVD-AMC.
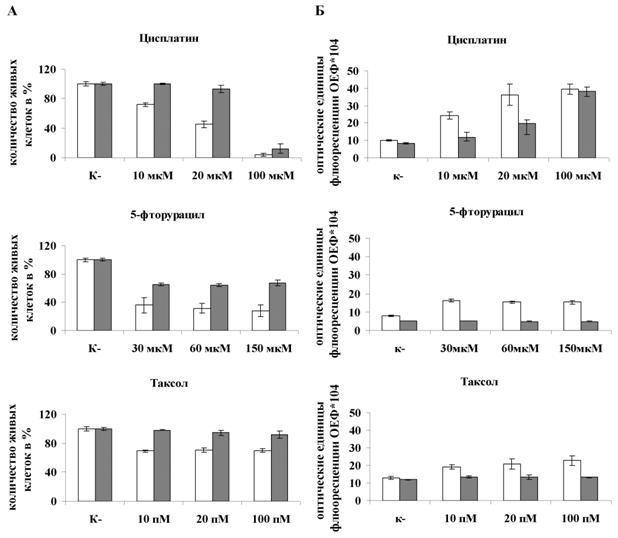
Рисунок 9. Выживаемость и активность каспаз 3/7 в клетках мышиной миеломоноцитарной лейкемии WEHI-3B при действии различных концентраций химиотерапевтических препаратов. (А.) Выживаемость клеток мышиной миеломоноцитарной лейкемии WEHI-3B; (Б) Активность каспазы-3/7. Данные по каждой точке являются результатом трех независимых экспериментов. (р<0,005)
Как видно из представленных результатов микоплазменная инфекция клеток WEHI-3B приводила к достоверному (р<0,005) повышению их выживаемости и снижению уровня активации каспаз-3/7 на 25-30% при различных внутриклеточных повреждениях по сравнению с неинфицированными клетками (белые столбцы).
Статистическую обработку результатов проводили общепринятыми методами с использованием программ Microsoft Excel и Statistica 6.0.
Таким образом, в первой части экспериментов in vitro было показано, что микоплазменная инфекция к опухолевым клеткам WEHI-3B, экспрессирующим набор Толл-подобных рецепторов 2 и 6, приводит к активации в них транскрипционного фактора NF-kB и подавлению апоптоза в данных клетках при различных внутриклеточных повреждениях. Полученные данные, были подтверждены в независимых экспериментах с использованием структурного компонента M.arginini – R-Pam2 (данные не приводятся). В экспериментах были использованы различные перевиваемые (B10M, 293TLR2/CD14, 293TLR2/6) и первичные (острый лимфобластный лейкоз, множественная миелома) культуры клеток, экспрессирующие специфические к микоплазме Толл-подобные рецепторы (см. главы 3.3.7 и 3.4. диссертации). Примечательно, что активация NF-kB в ответ на микоплазменную инфекцию (обработку структурными компонентами микоплазмы), существенно влияла на выживаемость клеток, но не влияла на скорость их пролиферации (см. главу 3.4).
Работа выполнена совместно с к. б.н. [лаборатория молекулярной биотехнологии ФГБУ «НИИЭМ им. » Минздравсоцразвития России].
Влияние микоплазменной инфекции клеток WEHI-3B на прогрессию мышиной миеломоноцитарной лейкемии в экспериментах in vivo
В следующей части работы было изучено влияние микоплазмы на прогрессию опухолевого роста в экспериментах in vivo.
Поскольку доступных иммунокомпетентных для M.arginini мышиных моделей нет, для проведения исследования использовали бестимусных мышей линии D2&I. Мышам вводили клетки WEHI-3B, инфицированные M.arginini в количестве 2х106 клеток/мышь. Каждая группа состояла их 8 мышей. Животных наблюдали в течение всего периода эксперимента до момента гибели последнего животного, регистрируя их общее состояние.
Анализ кривых выживаемости Каплана-Мейера (рис. 10А) показал достоверное (р<0,001 по лог-ранговому тесту) снижение продолжительности жизни мышей, с привитыми опухолевыми клетками WEHI-3B, инфицированными микоплазмой, по сравнению с контрольными животными, получавшими не инфицированные опухолевые клетки. Также, мыши в данной группе характеризовались более ранним развитием асцитов (рис. 10Б).
У мышей в группе с введением опухолевых клеток WEHI-3B, инфицированных M.arginini, развитие асцитов регистрировалось на 8 день после введения, в то время как у мышей, получавших неинфицированные клетки WEHI-3B, асциты регистрировались на 15 день после введения. Анализ проб асцитной жидкости показал, что в основном они были представлены опухолевыми клетками WEHI-3B, а также в пробах асцитов у мышей, получавших инфицированные опухолевые клетки, обнаруживали M.arginini в высоком титре.
Работа выполнена совместно с лабораторией д. м.н. [Институт Канцерогенеза РОНЦ РАМН]
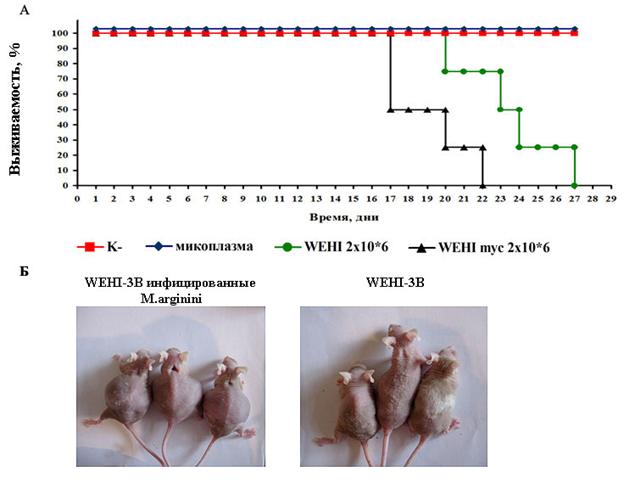
Рисунок 10. Влияние микоплазменной инфекции на прогрессию мышиной миеломоноцитарной лейкемии. (А.) диаграмма выживаемости бестимусных мышей линии D2&I. (р<0,001 по лог-ранговому тесту). (Б.) фотографии мышей линии бестимусных мышей линии D2&I.
Влияние диациллипопептида M. arginini на пролиферацию и резистентность к химиотерапевтическим препаратам опухолевых клеток WEHI-3B в экспериментах in vivo
В работах, выполненных и соавт., было показано, что при микоплазменной инфекции антигены микоплазм могут обнаруживаться в крови человека и животных в течение длительного периода. В данной части исследования было изучено влияние циркулирующих антигенов микоплазмы на пролиферацию и устойчивость к химиотерапевтическим препаратам клеток WEHI-3B.
На первом этапе нами была проведена оценка влияния диациллипопептидов на скорость прогрессии опухолевого роста. В эксперименте участвовало 40 животных (мыши линии Balb/C, самки) массой 18-20 грамм. Животные были разделены на 4 группы по 10 животных в каждой. Первую контрольную группу составили интактные мыши. Вторую группу составили мыши, которым трехкратно внутрибрюшинно вводили R-Pam2 (схема введения как в группе 4). Третью группу составили мыши, которым перевили клетки мышиной миеломоноцитарной лейкемии WEHI-3B в количестве 2*106 клеток/мышь. Четвертую группу составили мыши, которым внутрибрюшинно перевили клетки WEHI-3B в той же дозе. Этой же группе мышей через 1,3,5 суток после перевивания опухоли вводили R-Pam2 в количестве 5 мкг/мышь. Далее, для оценки прогрессии опухоли через 20 суток мышей безболезненно умервщляли с помощью диэтилового эфира и извлекали печень и селезенку. Отобранные органы использовались для макроскопического и гистологического исследования. Также был определен средний вес селезенки в каждой экспериментальной группе. (см. Рисунок 11 А-В). По данным макроскопического исследования, видно, что в группах 1 и 2 видимых патологических изменений не наблюдается (рисунок 11А.), но наблюдается незначительное увеличение среднего веса селезенки в группе 2 (рисунок 11Б.). При макроскопическом исследовании печени и селезенки мышей, которым был перевит промиелоцитарный лейкоз (группа 3), обнаружены характерные для лейкоза изменения: увеличение селезенки и некоторое увеличение печени. На поверхности печени и селезенки обнаружены редкие беловатые опухолевые образования. При макроскопическом исследовании печени и селезенки мышей, которым был перевит лейкоз и вводился диацилипопептид R-Pam2 (группа 4), также были обнаружены характерные для лейкоза изменения. Селезенки мышей были сильно увеличены. На поверхности селезенок и печени обнаружены беловатые рыхлые образования (которые при гистологическом исследовании оказались лейкозными миелоцитарными клетками).
Измерение средней массы селезенки показало, достоверное увеличение веса этого органа у лейкозных мышей (группа 3 и 4) по сравнению с контрольными группами 1 и 2. Введение R-Pam2 лейкозным мышам (группа 4) дополнительно увеличивало среднюю массу селезенки (p<0,05) по сравнению с аналогичным показателем для группы 3 (Рисунок 11Б.).
Гистологическое исследование селезенки и печени мышей из группы 1 и 2 патологических изменений не выявило. Гистологическое исследование селезенок мышей из групп 3 и 4 выявили сходную картину. В селезенках наблюдалась диффузная плотная инфильтрация пульпы лейкозными миелоцитарными элементами; лимфатические фолликулы были атрофированы. Наибольшие отличия обнаружены при изучении образцов печени мышей из групп 3 и 4. В печени мышей из группы 3 были отмечены многочисленные мелкие лейкозные миелоцитарные инфильтраты, тогда как в группе 4 инфильтраты были значительно крупнее (рисунок 11В.) Инфильтраты располагались преимущественно по ходу синусоидов. Скопления лейкозных клеток, также обнаруживалось в отдельных кровеносных сосудах. В группе 4, в отличие от группы 3 наблюдалась выраженная поверхностная инфильтрация печени лейкозными клетками (рис.11А).
Проведенное макро - и микроскопическое исследование образцов селезенки и печени мышей, которым внутрибрюшинно перевивали клетки мышиной миеломоноцитарной лейкемии WEHI-3B, позволило сделать заключение о том, что диациллипопептид микоплазмы положительно влияет на прогрессию опухолевого заболевания.
На следующей стадии мы провели дополнительный эксперимент по оценке влияние диациллипопептида R-Pam2 на скорость роста опухоли. При этом, одновременно, было оценено, каким образом R-Pam2 влияет на резистентность перевиваемых клеток к химиотерапевтическим препаратам. Эксперимент проводился по следующей схеме. Мышам линии BALB/c внутрибрюшинно прививали клетки мышиной миеломоноцитарной лейкемии WEHI-3B в количестве 2*106 клеток/мышь. Через 24 часа мышам системно вводили синтетический диациллипопептид M.arginini (R-Pam2) в дозе 5 мкг/мышь. Спустя двое суток мышам в течение 3 дней также вводили R-Pam2, а группы мышей с проведением химиотерапии получали еще и препарат 5-фторурацил в дозе 0,6 мг/мышь. В контрольные группы вошли животные, которым отдельно вводили R-Pam2 и 5-фторурацил. Каждая группа содержала 10 мышей. Животных наблюдали в течение всего периода эксперимента до момента гибели последнего животного (32 дня), регистрируя их общее состояние. На рисунке 11Г. представлена общая диаграмма выживаемости мышей, участвующих в эксперименте.
Анализ кривых выживаемости Каплана-Мейера показал, что мыши, получавшие синтетический диациллипопептид M.arginini, хуже поддавались лечению химиотерапевтическим препаратом 5-фторурацил по сравнению с мышами, не получавшими R-Pam2. В группе мышей с химиотерапией последняя мышь пала на 33 день, в то время как мыши, одновременно с химиотерапией получавшие R-Pam2, пали уже на 26 день.
Данные по выживаемости мышей в эксперименте in vivo полностью подтвердили полученные ранее результаты на культурах клеток. Более того, как видно из диаграммы, мыши в группе с одновременным введением клеток WEHI-3B и R-Pam2 полностью пали уже на 19 день, в то время как мыши, не получавшие R-Pam2 пали на 25 день. Полученные результаты свидетельствуют о том, что внутримышечное введение R-Pam2 приводило к повышению скорости прогрессии опухолевого роста и сокращению сроков жизни мышей. Примечательно, что полученные данные в описанном эксперименте не согласовывались с результатами на культуре клеток, где добавление R-Pam2 в культуральную среду не приводило к повышению кинетики роста клеток WEHI-3B.
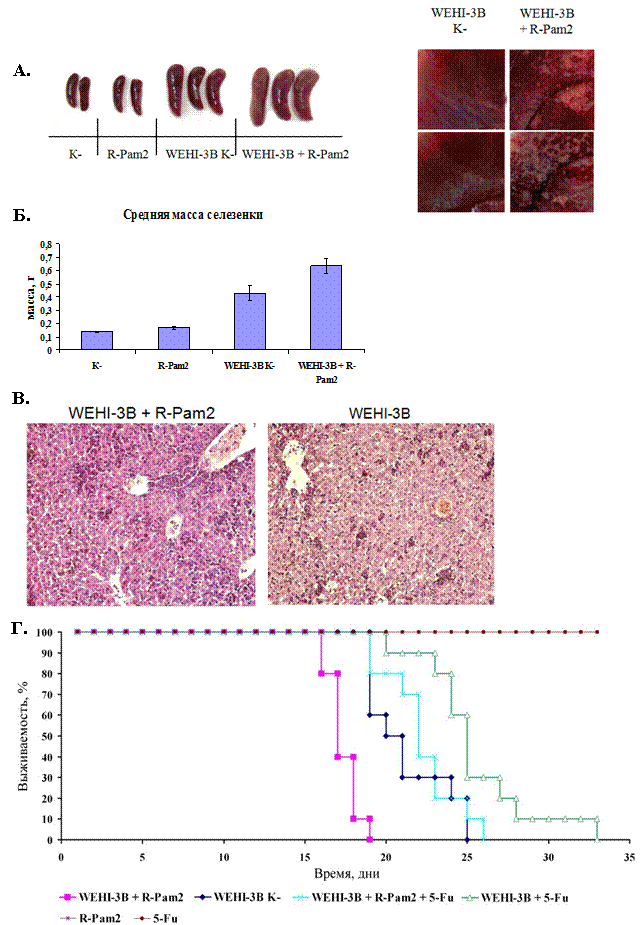
Рисунок 11. Влияние диациллипопептида M.arginini на пролиферацию и резистентность к химиотерапевтическим препаратам опухолевых клеток WEHI-3B
(А.) Макрофотографии органов мышей. Слева представлены макрофотографии селезенок, справа, участков печени с инфильтратами. K- группа интактных мышей, инъецированных ФСБ; R-Pam2- группа мышей, инъецированная диациллипопептидом R-Pam2; WEHI-3В группа мышей, с перевитыми опухолевыми клетками WEHI-3B; WEHI-3В+R-Pam2 - группа мышей, с перевитыми опухолевыми клетками WEHI-3B, инъецированная диациллипопептидом R-Pam2.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
