
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
(Б.) Инфекция клеток M. arginini снижает активность эктопически экспрессированного р53.
Инфицированные (черные столбцы) или неинфицированные (белые столбцы) р53-негативные клетки линии Н1299, содержащие р53-респонсивный элемент LacZ сеяли на 96-луночный планшет, после чего трансдуцировали рекомбинантными аденовирусами Ad-p53 (верхняя панель) или Ad-SEAP (нижняя панель) в дозе 5, 10 или 20 КОЕ на клетку. p53-зависимую трансактивацию репортерного гена и активность SEAP определяли через 24 часа после аденовирусной инфекции.
(В.) Изменение активности каспазы 3 в клетках инфицированных M.arginini.
Активность каспазы 3 определяли в лизатах клеток H1299 через 24 часа после инфекции (как описано в пункте Б.) при использовании флюорогенного субстрата.
(Г.) Инфекция M. arginini снижает рост-ингибирующий эффект эктопически экспрессированного р53.
Инфицированные и неинфицированные микоплазмой клетки были посеяны на 96-луночный планшет, после чего были трансфинфицированы рекомбиннатным аденовирусом Ad-p53 в количестве 5 КОЕ/клетка. Выживемость клеток оценивалась по их окраске метиленовым голубым в указанные интервалы времени.
(Д.) Эффект инфекции M. arginini на индукцию апоптоза в клетках E-Ras.
Клетки E-Ras-LXSP и E-Ras-Bcl2 были посеяны на 96-луночный планшет и инкубировались в бессывороточной среде (16 часов) или среде, содержщей 30 мкМ 5-ФУ(24 часа). Выживаемость клеток оценивалась по окраске метиленовым голубым (верхняя панель). Активность каспазы 3 определяли используя специфический флюорогенный субстрат (нижняя панель). Инфицированные M. аrginini клетки - черные столбцы, неинфицированные клетки - белые столбцы, клетки обработанные LAMPs (2 мг/мл за 4 часа до обработки 5-ФУ - серые столбцы.
инфекция не влияла на эффективность аденовирусной трансдукции и экспрессию целевого гена в составе генома аденовируса. При этом, напротив, из данных р53-зависимой трансактивации репортера (в клетках с эктопически экспрессированным р53), видно, что микоплазменная инфекция достоверно снижала активность р53 (Рисунок 3Б, верхняя панель). Эффект микоплазменной инфекции на индукцию р53-зависимого апоптоза был также оценен по активности каспазы 3 через 24 часа после трансдукции клеток аденовирусом. При всех использованных множественностях инфекции клеток Ad-p53, активность каспазы 3 была ниже в клетках инфицированных микоплазмой (Рис. 3В). Ингибирование р53-зависимого апоптоза в клетках инфицированных микоплазмой также было показано по изменению общего уровня выживаемости клеток Н1299, трансдуцированных Ad-p53 (см. Рис. 3Г).
Таким образом, полученные результаты показывают, что снижение транскрипционной активности р53 “транслируется” в ингибирование апоптоза - одной из важнейших р53-регулируемых функций.
Влияние микоплазменной инфекции на индукцию апоптоза было дополнительно исследовано с использованием клеточной линии E-Ras (линии крысиных фибробластов, трансформированных комбинацией Е1а и Ras). [Nelyudova А., 2004]. Были использованы два варианта этой клеточной линии: E-Ras-LXSP (клетки, трансдуцированные контрольным лентивирусом без вставки целевого гена) и E-Ras-Bcl2 (клетки, трансдуцированные ретровирусным вектором, обеспечивающим гиперэкспрессию антиапоптотического белка Bcl-2). Клетки E-Ras-LXSP являются высокочувствительными к различным проапоптотическим стимулам, тогда как клетки E-Ras-Bcl2 высоко резистентны к апоптозу, вследствие гиперэкспрессии Bcl-2.
В контрольных и инфицированных M. arginini клетках E-Ras-LXSP и E-Ras-Bcl2 был индуцирован апоптоз двумя независимыми способами (удалением сыворотки из ростовой среды или обработкой 5-ФУ). Спустя 24 часа была определена выживаемость клеток. Как показано на Рис. 15Д. (верхняя панель) микоплазменная инфекция приводила к повышению выживаемости клеток E-Ras-LXSP в ответ на оба способа индукции апоптоза. Апоптозингибирующее действие микоплазмы также было подтверждено по снижению активности каспазы 3 в инфицированных микоплазмой клетках (Рисунок 3Д, нижняя панель). Суммируя результаты данного эксперимента, можно заключить, что клетки E-Ras-LXSP инфицированные микоплазмой приобретали резистентность к апоптозу, сравнимую с клетками E-Ras-Bcl2.
Необходимо отметить, что снижение количества клеток E-Ras-Bcl2 в ответ на удаление сыворотки или обработку 5-ФУ связано с индукцией ареста роста, но не апоптоза [Nelyudova А., 2004]. Дополнительно, в данном эксперименте перед индукцией апоптоза мы инкубировали клетки с липид-ассоцииро-ванными белками (LAMPs) M. arginini. Однако нам не удалось показать, что структурные элементы микоплазмы способны блокировать развитие апоптоза в клетках E-Ras.
Подводя итог, можно сделать заключение о том, что микоплазменная инфекция способна эффективно супрессировать такие главные функции р53, как контроль клеточного цикла и регуляция апоптоза.
Работа проводилась совместно с к. б.н. [лаборатория молекулярной биотехнологии ФГБУ «НИИЭМ им. » Минздравсоцразвития России].
Микоплазменная инфекция способствует Ras-опосредованной трансформации крысиных эмбриональных фибробластов
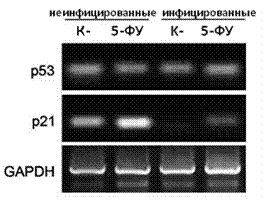
В клетках грызунов активность р53 является единственным фактором предотвращающим полную трансформацию клеток, экспрессирующих активированный онкоген Ras [Yaswen Р., 2007]. В связи с этим было интересно определить, является ли подавление р53, наблюдаемое при микоплазменной инфекции, достаточным для полной трансформации клеток, гиперэкспрессирующих онкоген Ras. Для проверки данного предположения в нашем исследовании была использована линия крысиных эмбриональных фибробластов REF52. Известно, что эти клетки характеризуются способностью к сенесенс-подобному аресту роста в ответ на экспрессию в них онкогена H-Ras. Инактивация р53 в этих клетках предотвращает арест и приводит к их злокачественной трансформации [Serrano М., 1997]. На первом этапе эксперимента мы показали, что микоплазменная инфекция приводит к снижению р53-зависимой транскрипции p21waf1 в клетках REF52 (Рисунок 4А.).Далее мы получили клеточную линию REF52 (REF52/sh-p53), в которой экспрессия р53 была подавлена в результате их трансфекции лентивирусом, экспрессирующим shРНК против р53 [Boiko А., 2006]. Нокдаун гена р53, опросредованный shРНК, был подтвержден полуколичественной ОТ-ПЦР по определению уровня мРНК р53. (Рисунок 4Б.). Далее клетки REF52 и REF52/sh-p53 были инфицированы микоплазмой M. arginini. После чего исходные неинфицированные клетки и клетки инфицированные микоплазмой были трансдуцированы лентивирусом (LV-Ras-Bleo), экспрессирующим онкоген Ras (H-Rasv12) в комбинации с геном устойчивости к блеомицину. Трансдуцированные клетки были помещены в среду с блеомицином для элиминации клеток нетрансдуцированных онкогеном Ras. Клетки были оставлены в этой среде на 3 недели для оценки изменения их морфологии и способности формировать колонии. Полученные результаты представлены на рисунке 4В. (слева, макро - и микрофотографии клеток, справа, схематическое изображение полученных результатов). Как и ожидалось через 21 день после селекции исходные клетки REF52 (содержащие функциональный р53 и неинфицированные микоплазмой) оказались арестованными и приобрели характерную морфологию для сененсентных клеток (Рисунок 4Б., панель ii). Клетки REF52/sh-p53 с нокдауном гена р53 и экспрессирующие онкоген Ras, напротив, сформировали полностью трансформированную культуру. (Рисунок 4Б., панель iii). При этом необходимо отметить, что клетки REF52/sh-p53, трансдуцированные онкогеном Ras, приобретали трансформированный фенотип вне зависимости от микоплазменной инфекции (неинфицированные клетки - панель vi, инфицированные - панель iii). Главным результатом данного эксперимента явилось то, что исходные (родительские) клетки REF52, инфицированные M.arginini, также формировали полностью трансформированную культуру, несмотря на тот факт, что они экспрессируют дикий тип гена р53 (панель v).
Таким образом, полученные результаты позволяют сделать заключение о том, что инфекция M.arginini, имитирует shРНК-опосредованное подавление активности p53, способствуя Ras-зависимой трансформации клеток REF52.
A. В.
Б.

Б.
Рисунок 4. Снижение активности р53 в инфицированных микоплазмой клетках способствует их Ras-опосредованной трансформации.А. РТ-ПЦР анализ экспрессии мРНК генов р53 и р21в клетках REF52, инфицированных и неинфицированных M.arginini.
Для индукции p53 клетки обрабатывали 5-ФУ (30 мкM) в течение 16 часов. Уровень экспрессии p53 и p21определяли методом РТ-ПЦР. Уровень экспрессия GAPDH (glyceraldehyde-3-phosphate dehydrogenase) был использован как позитивный контроль для подтверждения качества и количества кДНК в каждом образце.
(Б.) Ингибировние экспрессии мРНК p53 в клетках REF52, трансдуцированных shРНК против р53.
Уровень р53 был определен методом РТ-ПЦР через 72 часа трансдукции лентивирусом, экспрессирующим shРНК против р53. Экспрессия GAPDH (glyceraldehyde-3-phosphate dehydrogenase) была использована как позитивный контроль для подтверждения качества и количества кДНК в каждом образце.
(В.) Микрофотографии и фотографии зафиксированных и окрашенных метиленовым голубым клеток REF52.
Неинфицированные (панели i, ii и iii) и инфицированные M.arginini (панели iv, v и vi) клетки REF52 были трансдуцированы контрольным (без вставки целевого гена) лентивирусом (“K-“, панель i и iv) и лентивирусом LV-Ras-Bleo, экспрессирующим активированный онкоген Ha-Rasv12oncogene (панели ii и v) или комбинацией лентивируса LV-Ras-Bleo и лентивируса, экспрессирующего shРНК, специфичную к p53 (“shРНКp53”, панели iii и vi). Трансдуцированные клетки (за исключением отрицательного контроля, “K-“) культивировались в присутствии блеомицина в течение 21 дня. Выжившие клетки были зафиксированы и окрашены. Репрезентативные области сфотографированы при 200Х увеличении.
Изменение профиля экспрессии генов в клетках инфицированных микоплазмой.
Для более детального изучения влияния микоплазменной инфекции на функции р53 нами был проведен анализ изменения спектра экспрессии генов (около 35000 генов) в культуре клеток карциномы толстого кишечника человека HCT-116, зараженных и незараженных M.arginini. В анализ было включено 8 вариантов образцов:
А) Клетки HCT-116 с диким типом гена р53.
1) НСТ-116 (К-);
2) НСТ-116, обработанные 5-ФУ;
3) НСТ-116, инфицированные M.arginini;
4) НСТ-116, инфицированные M.arginini и обработанные 5-ФУ.
Б) Клетки HCT-116 с нокаутом гена р53(НСТ-116 р53-/-)
5) НСТ-116 р53-/- (К2-);
6) НСТ-116 р53-/-, обработанные 5-ФУ;
7) НСТ-116 р53-/-, инфицированные M.arginini;
8) НСТ-116 р53-/-, инфицированные M.arginini и обработанные 5-ФУ.
Перед определением профиля экспрессии клетки НСТ-116, с диким типом и нокаутом гена р53, зараженные и незараженные микоплазмой рассевали по 2х106 клеток на чашки диаметром 6 см. Через сутки половина клеток для активации р53 обрабатывалась 5-ФУ. Еще через 18 часов клетки собирали в Тризоле и экстрагировали из них РНК. Полученная РНК далее была использована для определения влияния микоплазменной инфекции на изменение профиля экспрессии генов (микроэррей-анализ проводился в подразделении Affymetrix при Институте рака Розвелла Парка из средств проф. ).
По результатам проведенного анализа были сформированы 3 основные группы генов (см. Табл№1): р53-зависимые гены, экспрессия которых подавляется микоплазмой; р53-независимые гены, экспрессия которых повышается в результате микоплазменной инфекции; р53-независимые гены, подавляемые микоплазмой. В представленную таблицу внесены гены, экспрессия которых изменялась наиболее значимо.
Таблица 1. Изменение профиля экспрессии генов в клетках инфицированных микоплазмой
Название гена | Изменение уровня экспрессии (разы) относительно клеток инфицированных мико-плазмой | |
р53-зависимые гены, экспрессия которых снижена в инфицированных микоплазмой клетках | - tumor protein p53 inducible protein 3 (TP53I3) - transformed 3T3 cell double minute 2, (MDM2) - BTG family, member 2 (BTG2) - cyclin-dependent kinase inhibitor 1A (p21, Cip1) - activating transcription factor 3 - TNF receptor superfamily, member 6 (Fas) | -1,55 -2,38 -1,53 -1,28 -1,62 -1,41 |
р53-независимые гены, экспрессия которых повышена в клетках инфицированных микоплазмой | - 5'-nucleotidase, ecto (CD73) - chemokine (C-C motif) ligand 26 - versican - tumor-associated calcium signal transducer 2 - calbindin 2, 29kDa (calretinin) - melanoma cell adhesion molecule - cystatin E/M - - GLIS family zinc finger 3 - alkaline phosphatase, placental - AXL receptor tyrosine kinase | 4.33 4.33 4.08 4.06 3.68 3.66 3.52 2.60 2.23 2.20 |
р53-независимые гены, экспрессия которых снижена в клетках инфицированных микоплазмой | - gap junction protein, alpha 1, 43kDa - myocyte enhancer factor 2C - methyltransferase like 7A - ras homolog gene family, member U - myocyte enhancer factor 2C - mannosidase, alpha, class 1A, member 1 - phosphodiesterase 4B, cAMP-specific killer cell - lectin-like receptor subfamily C, member - PDZ and LIM domain 5 | -3.96 -3.50 -3.37 -3.29 -2.76 -2.52 -2.36 -2.34 -2.30 |
р53-зависимые гены, экспрессия которых повышается в результате микоплазменной инфекции обнаружены не были. Гены, экспрессия которых регулируется 5-ФУ р53-независимым способом в таблицу не включены.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
