

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таблица 3. Размеры серотипоспецифических плазмид вирулентности
Серовар | Размер, п. н. |
S. gallinarum S. typhimurium S. enteritidis S. choleraesuis | 88230 93939 59372 49504 |
Помимо плазмид вирулентности встречаются крупные плазмиды до 200 килобаз. К таким плазмидам можно отнести группу коньюгативных плазмид IncHI размером 227000п. н. обеспечивающих утилизацию лактозы [24].
Плазмида S. abortusequi, размерами около 85 kb кодирует резистентность к токсичным тяжелым металлам (серебру, хрому, кадмию и мышьяку), у этой плазмиды также обнаружен ряд генов обеспечивающих резистентность к некоторым бета-лактамным антибиотикам и утилизацию цитрата [25].
Гены устойчивости к антибиотикам у сальмонелл кодируются как в хромосоме, так и некоторых плазмидах. Как правило, гены устойчивости к антибиотикам образуют кассеты, локализующиеся внутри транспозонов которые могут переноситься из хромосомы в плазмиду и обратно, учитывая коньюгативность большинства таких плазмид, этот механизм обеспечивает распространение генов антибиотикорезистентности среди сальмонелл [26]. Наиболее широко распространен у сальмонелл интегрон класса 1 состоящий из двух последовательностей – гена int I кодирующего интегразу и гена sul1 обеспечивающего устойчивость к сульфаниламидам. Данный интегрон встречается как в первом островке патогенности хромосомы, так и в плазмидах, причем благодаря активности интегразы, между генами intI и sul1 могут встраиваться другие гены антибиотикорезистентности [27, 28].
Процесс увеличения частоты встречаемости генетических детерминант антибиотикорезистентности носит обратимый характер и при снижении селективного давления частота плазмидо-кодируемых детерминант устойчивости снижается [29]. Среди R-плазмид ответственных за антибиотикорезистентность распространена плазмида класса IncFII 140 кб. несущая гены вирулентности spvABC и rck а также гены устойчивости к тетрациклину и хлорамфениколу [28]. Сходная в функциональном аспекте плазмида S. typhimurium относящаяся к IncFIC группе плазмид кодирует гены вирулентности spvCD вместе с детерминантами резистентности обеспечивающими реализацию ACSSuT фенотипа (устойчивость к ампициллину хлорамфениколу, стрептомицину, сульфаниламидам и тетрациклину) [30].
Среди функционально-значимых плазмид с низким молекулярным весом, найдено только несколько. Плазмида обеспечивающая перенос устойчивости к сульфаниламидам была описана у S. choleraesuis [31]. У S. typhi D4 мультикопийная плазмида кодирует систему рестрикции-модификации (RMS) [32]. Плазмида pWQ799 у серовара S. Borreze способна модифицировать О антиген [33]. Одной из важнейших функций низкомолекулярных плазмид кодирующих системы-модификации рестрикции может являться обеспечение фагорезистентности, в частности подобная роль показана для плазмиды pI, [14].
Таким образом, иммунохимическое и VNTR типирование сальмонелл абсолютно недостаточно для оценки патогенного потенциала полевых изолятов сальмонелл и адекватной оценки эпидемической значимости штаммов сальмонелл, обязательным условием молекулярно-эпидемиологического анализа сальмонеллезных инфекций должно являться выявление высокополиморфных хромосомных маркеров штамма и охарактеризация мобильных генетических детерминант патогенности

3.2 Выявление гена SpvC
Одним из ключевых факторов определяющих способность сальмонелл вызывать внекишечные формы сальмонеллеза у человека и животных является устойчивость к фагоцитозу.
Устойчивость сальмонелл к фагоцитозу обуславливается двумя процессами: кислотоустойчивостью и экспрессией генов Spv оперона. Продукты оперона Spv обеспечивают разрушение актина в цитоплазме клетки и таким образом предотвращают слияние фагосомы с лизосомой что и предотвращает гибель сальмонеллы внутри клетки. Таким образом, этот механизм позволяет сальмонеллам не только репродуцироватся в кишечнике, но и вызывать внекишечные инфекции.
Сальмонелл можно изолировать из различных объектов (пробы патологического материала, продукция животноводства и птицеводства, пробы кишечного содержимого). Для изоляции используют селективные и элективные среды (среда Эндо, селенитовый бульон, висмутсульфитагар). Культуры обладающие типичными для сальмонелл культуральными характеристиками дополнительно типируют на питательной среде урислелект 4 (BioRad) для исключения энтеробактеров, клебсиелл и протеев также растущих на висмутсульфитагаре. Идентификацию вида проводят с использованием биохимических тестов ПБДЭ «ДС Нижний Новгород» и real-time ПЦР.
Для выявления наличия Spv оперона у тестируемых культур используют ПЦР на ген Spv С. В реакции применяются праймеры со следующей структурой: `5-ACAGTGCTCGTTTACGACTTGAAT-3`, `5-TCACGCCCACTTCGTAGTTCA-3`. Реакцию проводят по следующей программе: линеаризация 95оС 30сек., отжиг праймеров 54оС 20сек, элонгация 72оС -40 сек, количество циклов – 40.
Результаты реакции учитывают путем электрофореза ПЦР продуктов в 6% полиакриламидном геле и после окрашивания бромистым этидием, в случае положительной реакции на электрофореграммах регистрировали ампликоны размером 553 п. н.(см. рис. 2)
 916 23 24
916 23 24
553пн
Рис. 2. Результаты ПЦР на наличие гена SpvC
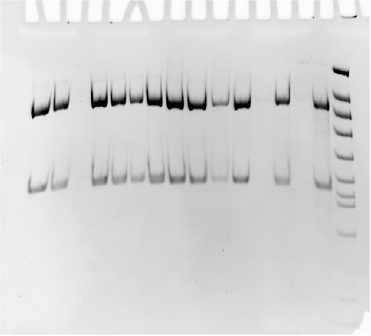 С целью повышения надежности результатов ПЦР, положительные на наличие гена SpvC пробы, следует дополнительно проверять путем рестрикции ПЦР-продукта эндонуклеазой Taq1. При этом образовывается 2 фрагмента длинной 395 и 158 п. н. (см. рис. 2)
С целью повышения надежности результатов ПЦР, положительные на наличие гена SpvC пробы, следует дополнительно проверять путем рестрикции ПЦР-продукта эндонуклеазой Taq1. При этом образовывается 2 фрагмента длинной 395 и 158 п. н. (см. рис. 2)
395п. н.
158п. н.
Рис.3 результаты верификации положительных на наличие гена SpvC проб рестрикцией Taq1.
Таблица 4. Результаты исследования наличия Spv региона у сальмонелл изолированных на с./х. предприятиях Сибири и Алтая.
наименование | Источник выделения | Кол-во культур имеющих SPV оперон | Количество тестированных культур |
Sl. typhymurium | Продукты животноводства | 2 | 2 |
Sl. choleraesuis | Печень птицы | 1 | 1 |
Sl. enteritidis | Печень птицы | 1 | 2 |
Sl. hamburg | ЖКТ птицы | 2 | 2 |
Sl. enterica | Органы яйцепродукции птиц | 3 | 6 |
Sl. dublin | Теленок, печень | 1 | 1 |
Всего культур протестировано | 14 | 42 |
Как следует из таблицы 4, около 33% культур имеют Spv оперон, что делает их опасными как в эпидемическом так и эпизоотическом отношении. Полученные результаты в целом совпадают с данными зарубежных исследователей [5,6].
Таким образом, система детекции и верификации высокопатогенных форм сальмонелл имеющих Spv оперон должна включать в себя спектр микробиологических техник обеспечивающих достаточную чувствительность и селективность, системы идентификации вида дублирующие друг друга но основанные на разных методических подходах и молекулярных механизмах. С целью повышения достоверности результатов ПЦР, исключения ложноположительных реакций не обусловленных кросс-контаминацией целесообразно ставить ПЦР только с плазмидной ДНК, а полученные положительные пробы дополнительно перепроверять рестрикцией.
3.3 Гены Rck и PagC
Salmonella могут проникать в нефагоцитирующие клетки благодаря наличию системы секреции III типа (T3SS-1), которая запускает триггер интернализации в клетку. Существуют данные что проникновение бактерий Salmonella enterica серовара Enteritidis в эукариотические клетки может также обеспечиваться внешним мембранным белком Rck
Rck – зависимая интернализация сальмонелл происходит путем активации комплекса Arp2/3 при участии малых GTPаз Rac1 и Cdc42. В результате формируется зиппер-подобное выпячивание мембраны эукариотической клетки обеспечивающее попадание сальмонелл внутрь клетки. Таким образом сальмонеллы являются единственными известными бактериями способными попадать внутрь клеток как через триггер-механизм, так и через зиппер-механизм интернализации [34].
Хотя у вирулентных штаммов сальмонелл наружние мембранные белки Rck and PagC имеют достаточно высокую гомологию по аминокислотным последовательностям (53%) сходство их функций не очевидно. Rck ассоциируется с устойчивостью к комплементу, адгезией на эпителии и инвазией [35, 36], тогда как PagC обеспечивает выживаемость при фагоцитозе и развитие системной инфекции [37]. Эти частичное расхождение функций лежит в основе участка с наибольшим сходством нуклеотидной последовательности локализованного в трансмембранной части токсинов, тогда как наибольший уровень различий нуклеотидных последовательностей наблюдается во внеклеточных доменах протеинов в основном определяющих межклеточные взаимодействия [38]. Тем не менее число сходных аминокислотных остатков больше между Rck и PagC чем между Rck , PagC и их ортологом Yersinia Ail, который подобно Rck обеспечивает адгезию и инвазию [39, 40]
3.4 Тестирование на наличие генов устойчивости к аминогликозидам StrA и StrB
StrA и StrB гены кодируют стрептомицин фосфотрансферразы Aph3 и Aph6 обеспечивающие устойчивость к стрептомицину но неактивные в отношении спектиномицина. Обычно эти гены локализуются внутри транспозона Tn5393 [41] и в «родительской» последовательности Tn5393c. [42, 43] свободной от IS элементов.
Sul2 ген кодирует сульфаниламид устойчивую дигидроптероат синтазу. Как отмечено ранее [42], RSF1010 и pSRC15 содержат протяженный 1778 п. н. сегмент транспозона Tn5393 который включает в свовем составе strAB и прилегающий к ним инвертированный повтор (IRR). Sul2 изначально не входили в состав этого генетического элемента, как следует из исследований некоторых авторов (GenBank последовательность. AB277723) Sul2 обнаружен в составе малого мобильного элемента CR2 [44]
п. н. сегмент транспозона Tn5393 который включает в свовем составе strAB и прилегающий к ним инвертированный повтор (IRR). Sul2 изначально не входили в состав этого генетического элемента, как следует из исследований некоторых авторов (GenBank последовательность. AB277723) Sul2 обнаружен в составе малого мобильного элемента CR2 [44]
3.5 Нитроцефиновый тест
-латамазы, группа ферментов разрушающих антибиотики лактамного ряда. Многие из этих ферментов кодируются в мобильных детерминантах патогенности (плазмидах и транспозонах) и могут передаваться горизонтально между бактериями одного вида, возможен также межвидовой перенос. Однако эффективность выявления устойчивости, связанной с продукцией БЛРС, с помощью традиционных методов оценки чувствительности остается крайне низкой. Все - лактамазы делятся на 4 типа A, B, C, D из них B тип является металлобетталактамазами и характеризуется широким спектром действия фермента. -лактамазы различаются также по спектру активности, наиболее опасными являются лактамазы расширенного спектра действия. Продукция -лактамаз расширенного спектра (БЛРС) – один из наиболее распространенных и клинически значимых механизмов резистентности энтеробактерий к современным -лактамным антибиотикам [45].
Одним из способов выявления бетталактамаз является нитроцефиновый тест, сущность теста заключается в выявлении бетта-лактамаз которые разрушают хромогенный субстрат – цефалоспориновый антибиотик цефиназу с образованием хромогенного субстрата. Для выявления нитроцефиновой (цефиназной) активности культур, следует ресуспендировать свежую культуру в капле физиологического раствора (1-2 бактериальных петли при отборе материала с твердой питательной среды или 1-2 капли бульонной культуры), и пропитать диск с нитроцефином. Нередко бетта-лактамазная активность выявляется сразу в течение 5-10 минут, в виде розового окрашивания, однако рекомендуется инкубировать диски при 37 С в течение 1 часа, желательно во влажной камере (например, чашке Петри). Особенность применения антибиотиков лактамного ряда, при терапии инфекций желудочно-кишечного тракта, равно как и при пероральном применении лактамных антибиотиков заключается в том, что защиту патогенного микроорганизма от антибиотика может обеспечить любой компонент микробиоценоза секретирующий фермент бетта-лактамазу. В связи с этим, мы рекомендуем тестировать нативное кишечное содержимое на наличие бетта-лактамазной активности.
Предложенная нами модификация метода заключается в следующем: свежий образец кишечного содержимого или фекалий (помета) отбирают ложкой Фолькмана или Люэра (предварительно обожженной на огне спиртовки). Образец ресуспендируют в 1 мл стерильного физиологического раствора, сначала путем смывания кишечного содержимого с ложки Фолькмана, потом с помощью пипетки. 50 мкл. суспензии наносят на предметное стекло в виде капли, аналогичным образом поступают с остальными пробами кишечного содержимого (ложку Фолькмана следует каждый раз очищать от остатков предыдущей пробы и обжигать над пламенем спиртовки). Затем на капли суспензий раскладывают диски с нитроцефином, предметное стекло помещают в чашку Петри и инкубируют не менее часа, после чего проводят учет результатов как описано выше (см. также рис.4). Метод удобен для обоснования назначения антибиотиков лактамного ряда а также мониторинга адаптации кишечных микробиоценозов к антибиотику.


Рисунок 4. Результаты тестирования проб помета на бетта-лактамазную активность (первые 4 пробы положительны, проба 5 - отрицательна).
4. Флюориметрические методы исследований сальмонелл для исследования фагоцитоза
Изучение фагоцитоза осуществляют с использованием свежей крови кролика и флюоресцентномеченных бактерий. Бактерии к заражению подготавливаются двумя различными способами:
1) В культуру клеток добавляют реакционно-способную группу (сукцинимидное производное флюоресцеина). Флюоресцентная метка связывается в щелочной среде рН 8 с лизином (положительно заряженная аминокислота) на поверхности прокариотической клетки.
2) К бактериям добавляют краситель флюоресцеина диацетат (ФДА) - его особенность — эффективный транспорт в живую бактериальную клетку. После его транспорта внутрь бактериальной клетки, эстераза отщепляет СН3 группу у ФДА и образуется флюоресцин флюоресцирующий в ультрафиолетовых лучах. Получается светящаяся изнутри бактерия.
Микроскопию проводят, например, с использованием эпифлюоресцентного микроскопа ImagerD1 (CarlZeiss) и программного обеспечения AxioVision. Также использовали конфокальный микроскоп фирмы Olympus.
Принцип метода состоит в том, что при фагоцитозе бактериальной клетки окрашенной снаружи, флюоресцеин может снижать свою флюоресценцию при закислении рН внутри фаголизосомы, а при использовании флюоресцеина находящегося внутри бактериальной клетки снижение светимости будет происходить только внутри погибших клеток.
Использование меченых бактерий позволит зафиксировать:
- образование фагосом
- этап слияния фагосомы с лизосомой — по снижению светимости клеток меченных сукцинимидным производным флюоресцеина снаружи и сохранению светимости живых бактериальных клеток меченных FDA.
Также в работе использовался краситель — трипановый синий, который гасит флюоресцентное свечение (квенчинг) у бактерий, которые не подверглись фагоцитозу.
Пробоподготовка окрашивания бактерий ФДА
1. Готовим предварительно рабочий раствор ФДА (из маточного раствора ФДА 1 мг/мл):
1мкл ФДА + 1мл Физиологического р-ра
В 4 пробирки вносим по 1 мл физ. раствора. Потом в них же в первую пробирку добавляем 1 мкл ФДА, во вторую 2 мкл ФДА, в третью – 3мкл, и в четвертую - 4 мкл соотвественно.
2. Готовим рабочую суспензию бактерий:
Использовали культуру клеток S.enterica Jl/bnc 13 = назовем ее A. В физ. растворе или PBS следует отмыть клетки от питательной среды. Для этого А — центрофугировали 3000 оборотов 2 минуты (далее 3000\2). Слили супернатант. Затем добавили 500 мкл PBS(рН=7,2 - 7,4) и центрифугировали 3000\2. Слили супернатант и опять добавили 500 мкл PBS, центрифугировали 3000\2, слили супернатант. Потом добавили 200 мкл PBS и разлили бактерии по четырем пипеткам по 50 мкл в каждую
3. Метим бактерии ФДА: для этого в суспензию бактерий капаем ФДА.
4. Инкубируем 30 минут.
5. Затем центрифугируя отмываем бактерии от непоглощенного флуоресцина (3-4 раза):
- центрифугируем 3000\2, сливаем надосодок
- +1000мкл физ р-ра и центрифугируем 3000\2, сливаем надосадок
- +1000мкл физ р-ра и центрифугируем 3000\2, сливаем надосадок
- +200мкл физ р-ра и смотрим на эпифлюоресцентном микроскопе через 10 минут, 20 минут, 1 час, 2 часа.
Пробоподготовки окрашивания бактерий сукцинимидным производным
Приготовление маточного раствора 2мг\мл = 2 мг сукцинимида + 1мл(1000 мкл) ацетона (необходим неполярный растворитель)
Сукц2 = Приготовление 0,2 мг/мл раствора = 450 мкл физ р-ра или дист Н2О + 50 мкл маточ. раствора (– хранится недолго из-за гидролиза).
Берем 6 пробирок и в каждую вносим 1,5 мл суспензии бактерий. Центрифугируем 5000/8, сливаем надосадок, добавляем в каждую пробирку 0,1М КБК 200 мкл (рН=8, готовим – 1 таблетка на 100 мл, по уже заданной концентрации), т. е. в каждой пробирке получаем 200 мкл смеси бактерий с КБК. Далее в первую пробирку добавляем 200 мкл сукцинимидной смеси, перемешиваем (плч 0,1 мг/мл). Потом берем 200 мкл из этой пробирки и вносим во вторую пробирку, перемешиваем(0,05 мг/мл). И т. д. со всеми пробирками. Потом все пробирки ставим на ночь в холодильник. Утром 2х кратное промывания в PBSе.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
