

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При такой идеальной схеме отбора, когда не учитывается браковка высокопродуктивных коров по болезням и другим причинам, ежегодный прирост удоя при сохранении условий кормления и содержания составит
Sd=3500кг-3000кг=500кг
Sэ=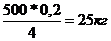
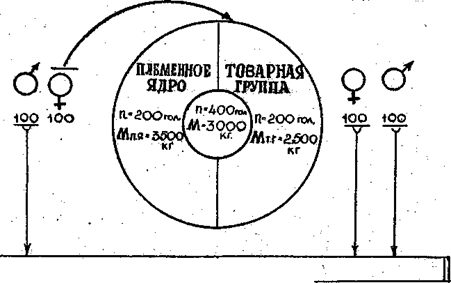 |
.ОТКОРМ
Рисунок 5
Однако выборка старых низкопродуктивных коров составляет значительно меньший удельный вес, чем в данной схеме. Селекционный эффект отбора в маточном стаде будет значительно меньше. Вместе с тем более существенную роль в повышении продуктивных качеств стада играют производители. За счет производителей можно создать более высокий генетический потенциал Sd. При одновременном учете влияния отбора и побора селекционный эффект вычисляют по формуле
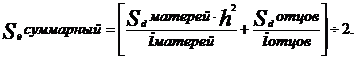
Если за стадом отделения закрепить быка – производителя - улучшателя, удой его дочерей превышает удой матерей на 200кг, следовательно, Sd отца =200кг.
Подставив данные в формулу получим
Sэ =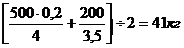
Предложенный метод расчета эффекта селекции не исчерпывает всех детерминант генетических изменений. Процессы, происходящие в популяции при отборе и подборе, значительно сложнее. Только творческий подход селекционера позволит получить поступательное приращение величины продуктивных качеств животных.
6 Генетическая структура популяции и анализ ее изменчивости
В животноводстве под популяцией понимают группу животных одной породы, имеющих фенотипические и генотипические различия и размножающихся путем свободного скрещивания друг с другом.
Для характеристики генетической структуры породы или стада, установления их родства и происхождения прибегают к изучению частот фенотипов, генотипов и отдельных аллелей, обуславливающих группы крови и полиморфные системы белков сыворотки крови, молока и лимфы тканей животных.
6.1 Определение частот фенотипов, генотипов и аллелей
Частота фенотипа в популяции - это относительное (процентное) количество особей, характеризующихся определенным фенотипом. Определяют частоту фенотипов по формуле
А=![]() ,
,
где А - частота определенного фенотипа в популяции;
n – число особей популяции определенного фенотипа(признака);
N- общее число особей в популяции.
Пример: среди стада симментальского скота учхоза «Прогресс» 112 коров красно-пестрой масти и 388 палево-пестрой. Необходимо вычислить частоту фенотипов красно-пестрых и палево-пестрых коров.
Частота красно-пестрых
А=![]()
Частота палево-пестрых
А=![]()
Сумма частот фенотипов должна быть рана единице или 100%.
Частота генотипа - это относительное число особей с определенными генами (АА, Аа, аа), выраженное в процентах или долях единицы от общего поголовья, составляющего целое (единицу и 100%).
Например, у КРС шортгорнской породы красная масть обусловлена генами АА, белая - аа, чалая – Аа. В данном случае расщеплению по фенотипу соответствует генотипам. В стаде, состоящем из N= 1610 голов, обнаружено 640 красных, 860 чалых и 150 белых животных. Отсюда
частота генотипа АА =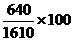 = 40 % (0,40);
= 40 % (0,40);
частота генотипа Аа = 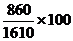 = 51 % (0,51);
= 51 % (0,51);
частота генотипа аа = 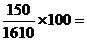 9 % (0,09).
9 % (0,09).
Зная частоты генотипов в популяции, можно установить частоты аллелей. При двухаллельной системе частоты аллелей вычисляют по формулам:
Частота аллели А рА=![]()
Частота аллели а qa= ![]()
Где р - частота аллели А;
q - частота аллели а;
n1 - число особей генотипа АА;
n2 - число особей генотипа аа;
n3 - число особей генотипа Аа;
N - общее число особей АА+Аа+аа.
При этом участвуют аллели А и А равна единице, т. е. р+q=1 (или 100%).
В тех случаях, когда гетерозиготы Аа фенотипические не отличаются от доминантных гомозигот АА, частоту генотипов аллелей вычисляют по формуле Гради - Вайнберга
р2+2рq+q2=1
(р2АА+2рq Аа+q2 аа=1)
Например, у кроликов белая окраска шерсти детерминированна геном а и является рецессивной по отношению к серой, контролируемой геном А.
В популяции из 200 животных 50 кроликов оказались белыми, остальные серыми. В данном случае генотипы АА и Аа не имеют фенотипических отличий, те и другие кролики будут серыми. Отсюда:
частота генотипа аа=![]() ;
;
частота аллели а=q=![]()
![]()
![]()
частота аллели А= р= 1-q=1-05=0,5;
частота генотипа АА=р2= 0,52=0,25;
частота генотипа Аа= 2рq= 2·0,5·0,5=0,5.
Выразив каждую величину в процентах, определяем количество животных в популяции с генотипами АА, Аа и аа. Так, доминантные гомозиготы АА составляют 25% от 200 голов, т. е. 50 кроликов, гетерозиготы Аа - 50% от 200 т. е. 100 кроликов.
6.2 Генетическое равновесие популяции
Генетическое равновесие популяции выражается формулой Гарди-Вайнберга. Это означает, что если в свободно размножающейся (панмиктической) популяции ген А встречается с частотой р, а его аллель а - с частотой q при р+q=1, то в пером же поколении устанавливается равновесие генотипов АА, Аа и аа.
Метод позволяет вычислить, соответствует ли фактическое расщепление фенотипов и генотипов любой популяции теоритически ожидаемому.
Например, изучая распространение безухости в популяции карапульских овец, по гену безухости установил следующие соотношения генотипов: 729АА+111Аа+4аа. Следует определить частоты аллелей фенотипов и генотипов и соответствие фактических генотипов теоретически ожидаемым по формуле Гарди-Вайнберга
Частота (фактическая) аллели А=р=![]() ;
;
Частота (фактическая) аллеля а=q=![]() ;
;
Частота генотипа АА=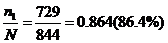 ;
;
Частота генотипа аа=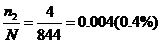 ;
;
Частота генотипа Аа=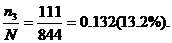
В популяции частоты генотипов совпадают с частотами фенотипов.
Расчет ожидаемых частот генотипов по формуле Гарди-Вайнберга начинают с рецессивных гомозигот. Если частота генотипа аа= q2=0,004, следовательно, частота аллея, а= √ q2=0,004=0,063. Отсюда частота аллели А=р=1- q=1-0,063=0,973.
Частота гомозигот А= р2=0,973=0,883.
Ожидаемые частоты генотипов будут следующими:
0,883 АА+ 0,113 Аа+0,004аа=1 или
88,3% АА+11,3% Аа+0,4% аа=100%,
а фактически частоты генотипов были
86,4% АА+13,2%Аа+0,4%аа=100% (844 гол.).
Таким образом, теоретически ожидаемые частоты генотипов близки к наблюдаемым.
Рассмотрим вычисление генетического равновесия на следующем примере.
У лошадей чистокровной верховой породы бывает носовое кровотечение, обусловленное рецессивным геном а. Его доминантный аллель А не вызывает носового кровотечения. На конном заводе среди 5400 голов 17 лошадей оказались с носовым кровотечением. Они погибли во время скачек. Необходимо определить, сколько животных в этой популяции являются носителями гена носового кровотечения, и каково вероятность появления лошадей с носовым кровотечением в следующем поколении при свободном спаривании фенотипические здоровых животных?
Частота генотипа 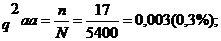
частота аллеля 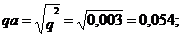
частота аллеля рА= 1- q=1-0,054=0,946;
частота генотипа рАА=0,9462=0,8,49%);
частота генотипа 2 рq Аа= 2*0,946*0,054=0,1,21%).
Следовательно, частоты генотипа по формуле Гарди-Вайнберга будут следующими:
0,8949 АА+0,1221 Аа+0,003 аа или
89,49% АА без носителя носового кровотечения (4832 гол.);
10,21% Аа с носителями гена носового кровотечения (551 гол.);
0,3% аа больных животных (17 гол.).
Если проводить элиминацию животных с генотипом аа, то структура популяции изменится. При этом число гетерозиготных животных Аа - носителей гена носового кровотечения - составил: n3= 0,1021*5400=551 гол. Количество гомозиготных животных будет:
n1=0,89499*5400=4832 гол.
Частоты при изменении состава генов определяем по формулам:
рА=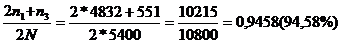 ,
,
р а=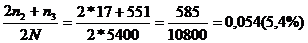
Отсюда появление животных с генотипом аа, составит 0,0029 или 0,29%, что на 0,01% ниже, чем в предыдущем поколении (таблица 8). Количество животных с носителем гена носового кровотечения снижается до 9,66% против 10,21%
Таблица 8
♀ ♂ | А= 0,9458 | А=0,054 |
А=0,9458 | АА=0,8945 | Аа=0,4830 |
а=0,054 | Аа=0,4830 | аа=0,0029 |
6.3 Определение генетического сходства популяций
При сравнении пород и популяций по сходству некоторых аллелей эритроцитарных антигенов крови или полиморфным белкам пользуются коэффициентом генетического сходства по формуле Майяла и Линдстрема
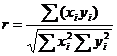 ,
,
где xi, yi - частоты одноименных аллелей в сравниваемых популяциях.
Чем больше r, тем больше степень генетического сходства популяций.
Например, по данным и частота встречаемости аллелей у черно-пестрого и голландского скота была следующей:
Таблица 9
популяция | Частота аллелей | |||||
A | F | Y | L | M | Z | |
Черно - пестрый скот, xi | 0,14 | 0,79 | 0,10 | 0,28 | 0,05 | 0,53 |
Голландский скот, yi | 0,16 | 0,80 | 0,07 | 0,13 | 0,02 | 0,40 |
Расчет данных для формулы производят по таблицы 10
Таблица 10
xi | yi | xi yi | xi2 | yi2 |
0,14 | 0,16 | 0,0224 | 0,0196 | 0,0256 |
0,79 | 0,80 | 0,6320 | 0,6241 | 0,6400 |
0,10 | 0,07 | 0,0070 | 0,0100 | 0,0049 |
0,28 | 0,13 | 0,0364 | 0,0784 | 0,0169 |
0,05 | 0,02 | 0,0010 | 0,0025 | 0,0004 |
0,53 | 0,40 | 0,2120 | 0,2809 | 0,1690 |
∑ xi yi=0,9108 ∑ xi2=1,0155 ∑ yi2=0,8478
Отсюда получаем:
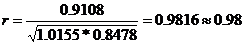 .
.
Сравниваемые популяции имеют большое генетическое сходство, что указывает на общность их происхождения.
7 Инбридинг. Вычисление коэффициента гомозиготности
Инбридинг - родственное спаривание животных. Инбридинг ведет к возрастанию гомозиготности потомков и разложению популяции на ряд генотипически различных линий, вследствие чего изменчивость инбридируемой популяции возрастает, а вариабельность каждой относительной линии снижается.
Инбридинг в разных формах используют в селекции, так как с его помощью можно выявить имеющиеся в популяции цепные комбинации генов и закрепить их в потомстве. Отрицательное последствия инбридинга - инбредная депрессия, ведущая к потере ценных комбинаций генов, имевшихся у гетерозиготных предков.
Степень инбридинга зависит от степени родства спариваемых животных. Определяют степень инбридинга по методу Рейта-Кисловского
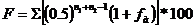 ,
,
где F - коэффициент инбридинга (степень гомозиготности);
0,5- доля наследственности одного родителя, передаваемая потомству; n1-ряд родословной, где встречается общий предок по материнской стороне; n2- ряд родословной, где встречается предок по отцовской стороне; fа - коэффициент инбридинга самого предка, если он инбридирован; ∑- знак суммы, указывает, что данные по каждому предку, на которого проведен инбридинг, суммированы.
Чем больше величина F, тем больше число генов у пробанда (потомка) переходит в гомозиготное состояние.
В зависимости от степени родства спариваемых животных различают инбридинг тесный (кровосмешение) F больше 25%;
Близкий =25,0…12,5%
Умеренный = 12,5…1,55%
Отдаленный меньше 1,55%
Гомозиготность пробанда можно оценить по его родословной (упорядоченной записи всех известных предков изучаемого животного на протяжении нескольких поколений).
Пример вычисления коэффициента гомозиготности для быка – производите6ля Рубина 119 дан в таблицы 11.
Таблица 11
М Лия | О Рыжик | ||||||||||||||
ММ Летопись | ОМ Надежда | МО Земляничка | ОО Нарядный | ||||||||||||
ММ Девчурка | ОММ Дубок | МОМ Малютка | ООМ Наполеон I | ММО Марта | ОМО Борис | МОО Неделя | I | ||||||||
ММММ Бонна | ОМММ Борис | МОММ Вишня | ООММ Борис | ММОМ Ева | ОМОМ Игрок | МООМ Дина | ОООМ Индус | МММО Неизвестная | ОММО Неизвестный | МОМО Неизвестная | ООМО Неизвестный | ММОО Венера | ОМОО Индус | М | О |
Степень родства III-III на Наполеона I и IY, IY-III на Бориса.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 |
