


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Обязательной структурой любой клетки является цитоплазматическая мембрана, которая отделяет цитоплазму от клеточной стенки. Толщина мембраны. 5-10 нм. При нарушении ее целостности клетки утрачивают жизнеспособность. Цитоплазма ряда бактерий пронизана мембранными структурами, которые являются производными цитоплазматической мембраны. У гетеротрофных бактерий их называют мезосомами. Они имеют вид пластинок (ламелл), пузырьков (везикул) или трубочек. Мезосомы могут быть расположены в зоне клеточного деления, вблизи нуклеотида и на периферии клетки, недалеко от цитоплазматической мембраны. У грамположительных бактерий мезосомальные структуры развиты в большей степени, чем у грамотрицательных. У фототрофных бактерий мембранные образования в виде пузырьков называют хроматофорами, а уплощенной формы - тилакоидами. Есть бактерии, у которых мембранная система не обнаруживается.
Определенную область в цитоплазме бактериальной клетки занимает нуклеоид. Он состоит из одной двойной спирально закрученной нити ДНК, замкнутой в кольцо. Ядерный аппарат прокариот не имеет ядрышка и не отделен от цитоплазмы мембраной. Через мезосомы нуклеоид связан с цитоплазматической мембраной. В период интенсивного деления в клетках ряда бактерий (Escherichia соli, Oscillatoria атоеnа) можно обнаружить несколько нуклеоидов.
В цитоплазме бактерий в свободном виде или в связи с мембранными структурами находятся рибосомы. Они имеют константу седиментации 70S, их размеры колеблются в пределах от 15 до 30 нм. Число рибосом может быть от 5 до 50 тыс., что зависит от возраста клетки и условий культивирования. Рибосом больше в молодых клетках.
В клетках различных бактерий часто обнаруживаются включения
запасных веществ. Это полисахариды, липиды, полифосфаты, сера. Они накапливаются при избытке тех или иных питательных веществ в окружающей среде, а расходуются при голодании. Из резервных полисахаридов особенно распространены глюканы: гликоген, крахмал и крахмалоподобное вещество - гранулёза. Они выявляются в клетках спорообразующих бактерий родов Bacillus и Сlоstгidium, а также у пурпурных бактерий и др. Полисахариды откладываются в цитоплазме равномерно или в виде гранул. Запасные липиды 6актерий представлены полиэфиром ![]() - оксимасляной кислоты и восками. Полиоксибутират накапливается на средах с избытком углерода у многих
- оксимасляной кислоты и восками. Полиоксибутират накапливается на средах с избытком углерода у многих
бактерий: видов Bacillus, Pseudoтanas, Spirilluт, Azotobacter, Sphaerotilus и др. Он обнаружен только у прокариот. Воска - эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий. Полисахариды и липиды служат хорошим источником углерода и энергии для клетки.
В условиях, препятствующих синтезу нуклеиновых кислот, у многих бактерий создается резерв фосфора в виде гранул полифосфатов. Впервые они были описаны у Spirillит volutans, поэтому их назвали волютином. Эти образования называют также метахроматиновыми зернами, так как они проявляют метахроматический эффект: приобретают красную окраску при обработке синим красителем.
Отдельные виды спорообразующих бактерий (Bacillus thuringiensis, Bacillus cereus, Bacillus popilliae и др.) в определенных условиях образуют в клетках кристаллы белковой природы, которые имеют правильную бипирамидальную форму и расположены непосредственно около споры. Их называют параспоральными тельцами.
Некоторые бактериальные структуры и включения, сильно преломляющие свет (эндоспоры, аэросомы, отложения полиоксибутирата и серы), хорошо заметны в световом микроскопе без специальной обработки. Часть структур (жгутики, клеточная стенка, нуклеоид, волютин и др.) можно выявить с помощью светооптического микроскопа только после окрашивания соответствующими красителями. Ряд структурных элементов бактерий - микрокапсулы, ворсинки, мезосомы, рибосомы и др. различимы только 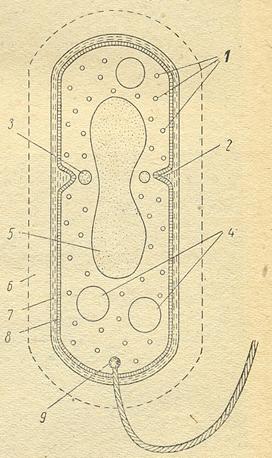
Рисунок 16. Схема строения бактериальной клетки: 1 – рибосомы, 2 – начавшееся образование поперечной перегородки, 3 и 4 – запасные отложения, 5 – ядерный район, 6 – капсула, 7 – стенки клетки, 8 – протоплазматическая мембрана, 9 – зерно, от которого начинается жгутик
в электронном микроскопе (рис.16).
ДРОЖЖИ
В группу дрожжей объединяются грибные организмы, которые в ростовой фазе существуют преимущественно в виде отдельных клеток. Примерно 2/3 дрожжей относится к классу сумчатых грибов(Ascoтycetes). Дрожжи есть также в классе базидиомицетов (Basidioтycetes) и дейтеромицетов - несовершенных грибов (Deuteroтycetes - Fungi iтperfecti).
Клетки разных видов дрожжей морфологически весьма разнообразны. Они бывают круглые, овальные, цилиндрические, яйцевидные, лимоновидные, колбовидные, треугольные, стреловидные и серповидные. Дрожжевые клетки значительно крупнее бактериальных. Длина их варьирует у разных видов от 2 до 20, иногда до 50 мкм, ширина от 1,5 до 10 мкм. К числу наиболее крупных дрожжей относятся, например, представители родов Saccharoтyces и Lipoтyces, наиболее мелкими являются виды Pichia и Наnsenula.
Как уже отмечалось, дрожжи существуют в основном в одноклеточной форме. Однако в культурах многих видов наблюдаются скопления разных размеров и очертаний, а некоторые дрожжи на определенной стадии развития могут образовывать мицелиальные структуры (рис. 17). Дрожжи являются неподвижными организмами.
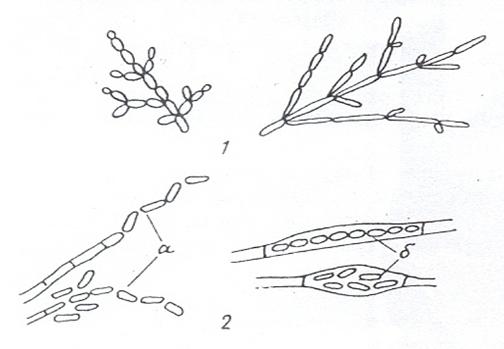
Рисунок 17. Мицелиальные формы дрожжей: 1 – псевдомицелий; 2 – истинный мицелий; а – артроспоры; б – эндоспоры
Дрожжи размножаются разными способами: вегетативно, бесполыми спорами и половым путем. Наиболее распространенным способом вегетативного размножения является почкование (рис. 18, 1). К почкующимся дрожжам относятся, например, виды родов Saccharoтyces и Lipoтyces. Если при почковании вновь возникающие клетки не отделяются друг от друга, то образуется псевдомицелий. Он отличается тем, что в зоне перегородок между клетками имеются перетяжки (рис. 18, 1). Образование псевдомицелия особенно характерно для дрожжей рода Candida.
Размножение делением (рис. 18, 2) встречается реже. Оно свойственно, например, видам Schizosaccharoтyces. Когда не расходятся делящиеся клетки, формируется истинный мицелий. Между клетками истинного мицелия четко различается перегородка, но нет перетяжек (рис. 18, 2). У ряда дрожжей (определенные виды Saccharoтyces, виды Nadsonia и др.) наблюдается так называемое почкующееся деление, при котором почка формируется на широком основании (рис. 18, 3). Почкующееся деление имеет морфологическое сходство с почкованием и делением.
Рисунок 18. Способы вегетативного размножения дрожжей: 1 – почкование; 2 – деление; 3 – почкующееся деление
Бесполое и половое размножение связано с формированием специализированных репродуктивных структур. При половом размножении их появлению предшествует слияние клеток и последующее объединение ядер, при бесполом размножении предварительное слияние клеток и ядер не происходит.
Размножение бесполыми спорами - баллистоспорами - характерно для видов Sporoboloтyces, Sporidiobolus и др. Баллистоспоры возникают на заостренных выростах клеток - стеригмах (рис. 19). При созревании они с силой отбрасываются на некоторое расстояние. Баллистоспоровые дрожжи относятся к базидиомицетам. Истинный мицелий таких дрожжей может распадаться на 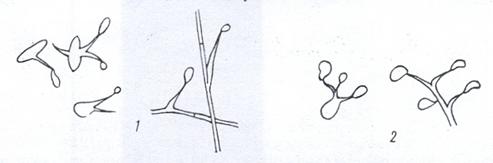
Рисунок 19. Баллистоспоры на отдельных клетках и гифах мицелия: 1 – Sporobolomyces; 2 - Sporidiobolus
отдельные клетки, называемые артроспорами.
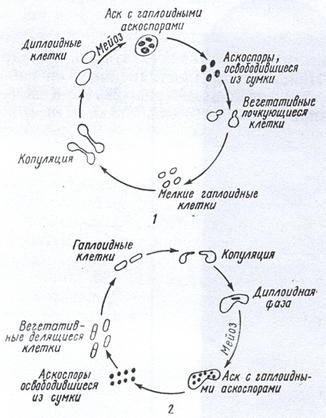
Рисунок 20. Жизненные циклы некоторых дрожжей: 1 – Saccharomyces cerevisiae; 2 – Schizosaccharomyces (Octosporomyces) octosporus
Половое размножение большинства дрожжей (аскомицетов) связано с образованием асков (сумок) и аскоспор. у одних (виды Saccharoтyces, Debaryoтyces и др.) образование асков происходит сразу после изо - или гетерогамной копуляции почек или вегетативных гаплоидных клеток в диплоидную. У других (виды Saccharoтyces, Schizosaccharoтyces, Saccharoтycodes и др.). Аски формируются из ранее возникших диплоидных клеток партеногенетически, т. е. без непосредственно предшествующей копуляции. В асках в результате мейоза образуются гаплоидные аскоспоры. В каждом аске бывает, как правило, от 2 до 8 спор. Иногда (виды Liротуces) их число достигает нескольких десятков. Аскоспоры многих дрожжей (виды Saccharoтyces, Schizosaccharoтyces) освобождаются из сумок и превращаются в вегетативные клетки ( рис.20).
У базидиомицетов в результате полового процесса развиваются специализированные репродуктивные клетки - базидии, на которых образуются экзогенные споры (базидиоспоры).
Обычно половой процесс чередуется у дрожжей с вегетативным размножением. Однако длительность гаплоидной и диплоидной фаз у разных дрожжей неодипакова. Например, У Sассharотусеs cerevisiae преобладает диплоидное состояние, у Schizosaccharoтyces (Octosporomyces) роmbе - гаплоидное. Кроме того, половой процесс и образование аскоспор у дрожжей происходит только в определенных условиях. В культурах Saccharoтyces активное спорообразование можно наблюдать, если молодые активные клетки, выращенные на полноценной богатой среде, поместить в условия недостаточного питания, плохого снабжения кислородом и влагой..
К половому процессу способны многие дрожжи. Но есть и такие, у которых половой процесс и спорообразование не обнаружены. Их включают в класс несовершенных грибов. К ним относятся представители Caпdida, Torulopsis и Cryptococcus.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
