

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
После заселения дождевыми червями разложение отходов ускоряется. В лабораторных опытах минерализация органического вещества активного ила протекала в 2-5 раз активнее, если черви присутствовали в нем. Однако, такая экстремально высокая скорость разложения возможна лишь при очень высокой плотности червей - до 20 г сырой биомассы/100 г осадков (Loehr et al., 1985). Это соответствует 40-60 червям в 100 г. Целлюлоза - необходимый компонент диеты дождевого червя, несмотря на то, что ее химическое разложение в процессе вермикомпостирования незначительно отличается от такого для обычного компоста (Hartenstein, 1983; Hatanaka et al., 1983; Hand et al., 1988). Целлюлоза нужна червям как источник пищевых волокон. Тем не менее, содержание органического углерода в органическом веществе копролитов ниже, чем в исходном субстрате, а степень его окисленности выше (Hatanaka et al., 1983; Grappelli et al., 1987). В вермикомпосте по сравнению с обычным компостом также падают соотношение C:N и потери при прокаливании (Satchell, 1983), возрастает электропроводность водного раствора (Hand et al., 1988). Когда соотношение органическое вещество/зола падает до значения 2, скорость воспроизводства червей снижается (Hatanaka et al., 1983). Возможно, этот показатель целесообразно использовать для определения степени зрелости вермикомпоста.
Дождевые черви способствуют переводу химических элементов в доступную для растений форму. Однако, в чем суть этого "посредничества" не известно. Копролиты содержат в 5 раз больше доступного азота, в 7 раз больше доступного калия и в 1,5 раза больше кальция, чем в среднем содержится в 15-ти см слое плодородной почвы. Время жизни питательных элементов в вермикомпостах в 6 раз больше (до 30 суток), чем в окультуренных почвах. Плодородные свойства вермикомпоста примерно в 5 раз выше, чем хорошо окультуренной почвы (Edwards, Fletcher, 1988).
Многие авторы отмечают относительное накопление в копролитах гумусированных химических структур, переходящих в щелочные вытяжки и придающих темную окраску последним (Курчева, 1971; Козловская, 1976). Однако механизм гумусообразования в присутствии дождевых червей не расшифрован. Очевидно, что основные химические события в вермикомпостах, связанные с дождевыми червями, следует искать на уровне биохимической трансформации, опосредованной микроорганизмами.
2. Взаимодействие двупарноногих многоножек и микроорганизмов
Диплоподы - одни из основных активных первичных разрушителей лестной подстилки. Они связанны с лесными почвами в разных природных зонах и перерабатывают опад и древесину. Таким образом, они являются одними из ключевых сапрофагов почвы. Среди пищевых энзимов у них отмечена высокая активность карбогидраз, расщепляющих олигосахара, протеаз и липаз (Nielsen., 1962; Marcuzzi, Turchetto., 1976).
В основе взаимодействия микроорганизмов и кивсяков лежат те же принципы, что и для взаимодействия их с дождевыми червями.
1. Это осуществление отбора микробных популяций и модификация микробных сообществ за счет пассажа пищевых субстратов через свой пищеварительный тракт. Переваривание микроорганизмов животными основано на "киллерном" эффекте. Есть такая работа по изучению киллерной активности пищеварительной жидкости кивсяков Pachyiulus flavipes, где подробно показан этот механизм (Бызов., 2005)
2. Не стоит забывать, что кишечник кивсякой также специфическое местообитание в почве, как и у кишечник дождевых червей, отличающееся по составу и активности микробов. Функциональная роль кишечных симбионтов беспозвоночных во многом аналогична таковой у позвоночных. Особенностью и отличием от дождевых червей является наличие в кишечнике кивсяков перитрофической мембраны, расположенной в средней кишке. Она создает барьер, который бактерии не могут преодолеть без дополнительного фактора-стрессора.
3. Регуляция по трофическому механизму основана на мобилизации животными питательных веществ, связанных в микробных клетках, их экскреции и высвобождению в почву микроорганизмов, размножающихся в кишечнике. Регуляция по метаболическому механизму предполагает физиологический отклик микробов на метаболиты животных, обладающих свойствами физиологически активных веществ. Предполагается, что у кивсяков есть свой энзим, действие которого многократно усиливает активность микрофлоры в пищевом комке (Стриганова., 1980).
Метаболический механизм объясняет быструю реакцию микробов на воздействие животных, не пропорциональную количеству потребленной биомассы. Обилие бактерий в кишечники кивсяков на примере Pachyiulus flavipes составляет 109-1010 КОЕ в экскрементах и 107 КОЕ в корме. (Бызов, 2003).
3. Основы кинетики микробного роста
Параметры роста и его анализ
Все подходы к изучению микробных организмов напрямую зависят от задач, которые ставятся перед исследователем. Так, например, для многих из них достаточно качественной характеристики роста, т. е. происходит ли он в культуре или же нет. Количественно, рост обычно изображают в виде графика зависимости биомассы от времени. Существуют различные параметры роста, такие как удельная скорость роста, время удвоения биомассы, лаг-период, или фаза задержки роста, метаболический коэффициент, экономический коэффициент. Рассмотрим некоторое из них.
Удельная скорость роста.
Для роста биомассы необходимы следующие условия:
Формула удельной скорости роста выглядит следующим образом
dx/dt= µx, где dx увеличение биомассы, за бесконечно малый промежуток времени dt, х - обилие (это может быть количество клеток, биомасса одной особи, КОЕ и др), а µ - удельная скорость роста, измеряется в единицах обратных единицам времени (1/t).
Интегрируя получаем следующие уравнения:
ln x = ln x0 +µt
ln( x/x0)=µt
x=x0eµt
Рост подчиняющийся этому закону, называется экспоненциальным или логарифмическим. Необходимо отметить, что данный закон выполняется, если условия окружающей среды и состав биомассы остаются постоянными. Удельная скорость роста является основным параметром, характеризующим скорость роста.
Выделяют так же время удвоения биомассы, степень размножения, обратное время удвоения, вычисляемые по уравнениям соответственно:
1) td=ln2/µ=0.693/µ
2) x/x0=2n => n= 3.32 log(x/x0)
3) 1/td или log2x=log2x0 + t/td
Экономический коэффициент.
Определяется уравнением:
Дx/Дs=Y, где Дx - увеличением биомассы, соответствующее потреблению субстрата в количестве Дs, Y - экономический коэффициент.
Или же в дифференциальной форме, при ds стремящемся к 0 , выглядит следующим образом: Y = - dx/ds. Знак минус вводится по тому, что x и s изменяются в разные стороны, а коэффициент принято считать положительным числом. Важность данного параметра состоит в том, что он выражает количественные потребности организма в пище.
Метаболический коэффициент.
Скорость потребления субстрата культурой в данный момент времени. Выражается следующим уравнением: ds/dt= qx, где ds - потребление субстрата, dt - момент времени, x - биомасса, q – метаболический
коэффициент или удельная скорость метаболизма.
Иначе можно записать, как µ / Y = q
Влияние концентрации субстрата на скорость роста.
Существование фазы экспоненциального роста в периодической культуре доказывает, что скорость роста может в довольно широких пределах не зависеть от концентрации субстрата. Следовательно, рост в этом случае характеризуется кинетикой нулевого порядка. Отсюда появляется предположение, что кинетике потребления субстрата будет соответствовать кинетика ферментативной реакции.
q=qms/(s+Ks) , где s - концентрация субстрата, q – метаболический коэффициент, qm - максимальное значение q, полученное при s>>Ks, Ks - константа насыщения, эквивалентная константе Михаэлиса-Ментен. Данные по определению Ks немногочисленны, поскольку значения Ks крайне мыла, часто ниже чувствительности химических методов определения. Численно ровна количеству субстрата S соответствующего µmax/2.
Лаг-период.
Максимальному уровню удельной скорости роста часто предшествует лаг-период. Длительность лаг-периода (![]() ) для многих целей удобно описывать методом Лоджа и Хиншельвуда. Если на графике зависимости логарифма биомассы от времени прямую линию экстраполировать до уровня начальной биомассы, то отрезок, отсекаемый при этом на оси абсцисс, и будет соответствовать
) для многих целей удобно описывать методом Лоджа и Хиншельвуда. Если на графике зависимости логарифма биомассы от времени прямую линию экстраполировать до уровня начальной биомассы, то отрезок, отсекаемый при этом на оси абсцисс, и будет соответствовать ![]() . Тогда уравнение для роста культур приобретает вид:
. Тогда уравнение для роста культур приобретает вид: 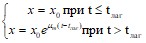
Предельные границы максимальной концентрации биомассы.
Существует понятие о максимальной плотности биомассы, которая может быть достигнута в данной среде. Она определяется одним из четырёх условий:
1) количество лимитирующего субстрата в среде
2) накопление ингибирующего продукта
3) максимальная плотность упаковки биомассы
4) отмирание клеток
Высказано несколько предположений. Например, Бейл говорил, что биологическое пространство значительно шире физического пространства, занимаемого организмом. На данный момент эта концепция отвергнута. Таким образом максимальная плотность упаковки клеток (число клеток в 1 см3) составляет около 1012/V, где V - объем индивидуальной клетки в мкм2
Биомасса и её определение.
Биомасса - общий термин, используемый для обозначения организмов в культуре. Существует несколько методов измерения биомассы. Сухая биомасса. Прямое определение количества сухой биомассы включает в себя отделение организма от среды, отмывание и высушивание его в печи при температуре 105 +/- 2 0C. Недостаток в том, что помимо биомассы среда не должна содержать неизвестное количество каких-либо твёрдых веществ.
Сырая биомасса. Проводят путем прямого взвешивания, опуская этап высушивания. Недостаток - не так точен, как метод определения сухой массы.
Объем. Количество биомассы можно сравнивать, измеряя их объемы, т. к. по плотности биомассы различаются незначительно.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
