

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
или в виде одного интегрального уравнения:
![]() , где
, где
мm – максимальная удельная скорость роста (1/ч)
х(t) – концентрация клеток (КОЕ/мл) в момент времени t (ч)
r0 – начальное значение переменной физиологического состояния r
x0 – начальная концентрация клеток бактерий в момент инокуляции (КОЕ/мл).
В модели X (концентрация клеток в среде), X0 (исходная концентрация в суспензии культивируемых бактерий) и t (время) – экспериментальные данные, а мm и r0 параметры. мm - максимальная удельная скорость роста. r0 - функция физиологического состояние растущей культуры, отражающее отличие удельная скорость роста м от мm в лаг-фазе. Параметр r0 характеризует физиологическое состояние в момент интродукции (в почве). На основании этого параметра вводится метаболическая готовность к росту. Под метаболической готовностью к росту – это фактически готовность организма к росту на определенной среде. Мы полагаем, что метаболическая готовность к росту определяется физиологическим состоянием. r0 характеризуется колебаниями на несколько порядков в связи с чем мы считаем необходимым логарифмирование этой величины. А для создания прямой зависимости (чем этот параметр выше, тем метаболическая готовность к росту выше) необходимо добавить заведомо большое число. Таким образом ![]() .
.
3. Описание эксперимента
Дождевые черви вида Aporrectodea caliginosa собираются на границе «новой» Москвы и Московской области на залежи 20 лет. Черви содержатся неделю на образце гумусо-аккумулятивного горизонта … почвы в условиях лабораторного микрокосма при постоянной влажности 60%. Далее черви отмываются от почвы, помещаются в чашку Петри и выдерживаются сутки при температуре +10°. После чего отбираются копролиты и вместе с почвой, в которой содержаться черви (корм), микробиологически анализируются.
Кивсяки вида Сilindroiulus caeroleocinctus собираются в городской подстилке Москвы в парке. Содержатся на подстилке взятом на месте отбора в условиях микрокосма. Далее отбираются экскременты червей и образца опада и миробиологически анаизируются.
4. Результаты
Количественный посев из ячеек планшеты позволяет установить зависимость между оптической плотностью и концентрацией клеток в суспензии. Подобная зависимость позволяется изучать рост периодической культуры по оптической плотности.
Корреляционный анализ показывает, что наблюдается прямая линейная зависимость оптической плотности от концентрации бактерий в суспензии (рис.2). Уравнение пересчета оптической плотности в концентрацию клеток: x=aЧOD, где x – концентрация клеток в суспензии (КОЕ/мл), OD – оптическая плотность при 620нм, a = 2,7±0,6Ч109 – коэффициент пересчета.
r=0,77 – коэффициент корреляции, r2=0,59 – коэффициент детерминации
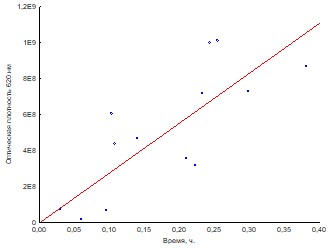
Рис. 2. Зависимость оптической плотности от концентрации бактерий в суспензии
Исходные данные:
Кривые периодического роста ассоциаций бактерий на жидких средах содержащих различные биополимеры, представленные на рисунках 3-10 для дождевых червей и с 11-20 для кивсяков.
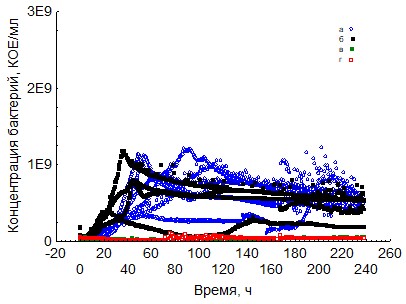
Рис. 3. Кривые периодического роста ассоциаций бактерий на крахмале (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
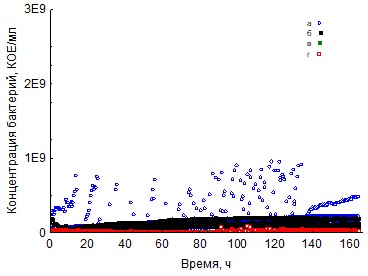
Рис. 4. Кривые периодического роста ассоциаций бактерий на КМЦ (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
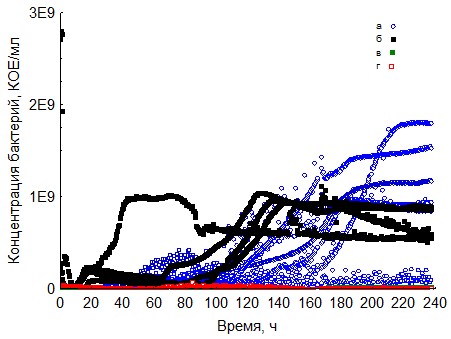
Рис.5. Кривые периодического роста ассоциаций бактерий на пектине (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
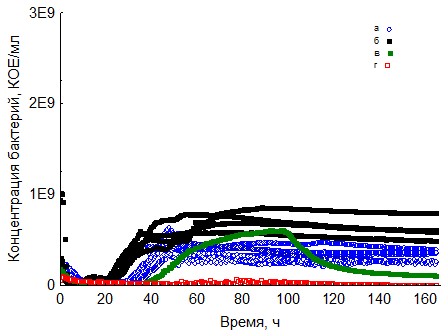
Рис. 6. Кривые периодического роста ассоциаций бактерий на ксилане (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
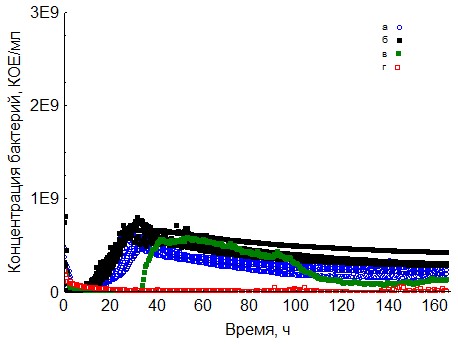
Рис. 7. Кривые периодического роста ассоциаций бактерий на казеине (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
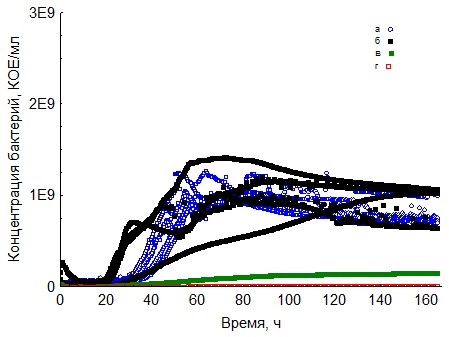
Рис. 8. Кривые периодического роста ассоциаций бактерий на декстране-500 (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
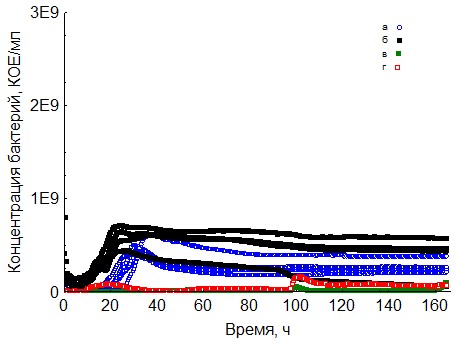
Рис. 9. Кривые периодического роста ассоциаций бактерий на твин-20 (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества)
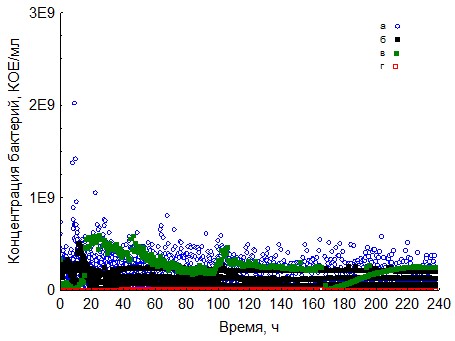
Рис. 10. Кривые периодического роста ассоциаций бактерий на хитине (a - корм, б - копролиты, в - контроль на стерильность, г - контроль на рост за счёт собственно органического вещества).
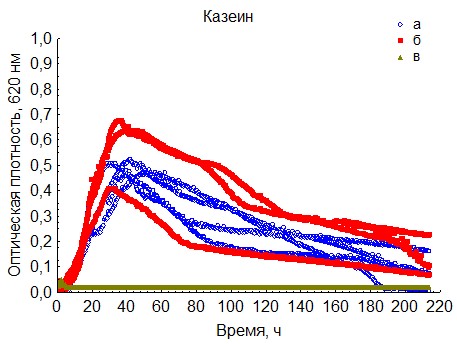
Рис. 11. Кривые периодического роста ассоциаций бактерий на казеине (a - корм, б - экскременты, в - контроль на стерильность).
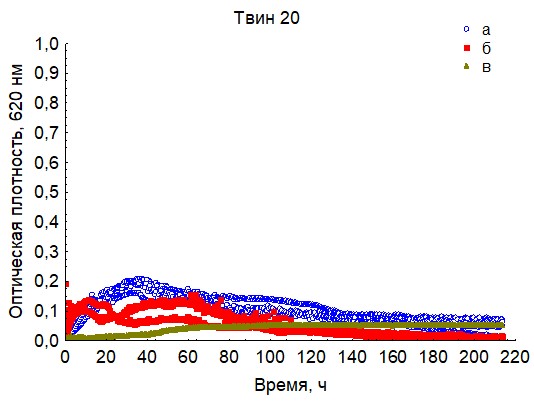
Рис. 12. Кривые периодического роста ассоциаций бактерий на твине 20 (a - корм, б - экскременты, в - контроль на стерильность).
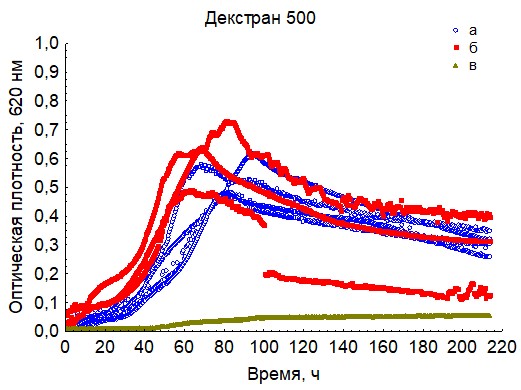
Рис. 13. Кривые периодического роста ассоциаций бактерий на декстран 500 (a - корм, б - экскременты, в - контроль на стерильность).
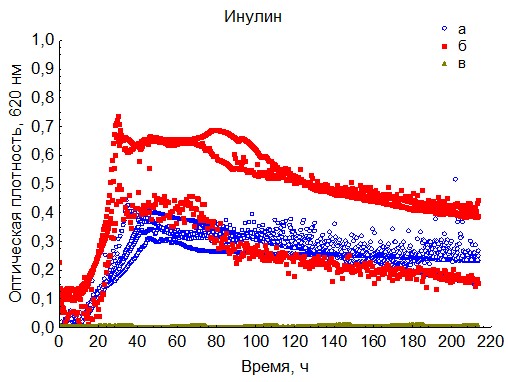
Рис. 14. Кривые периодического роста ассоциаций бактерий на инулине (a - корм, б - экскременты, в - контроль на стерильность).
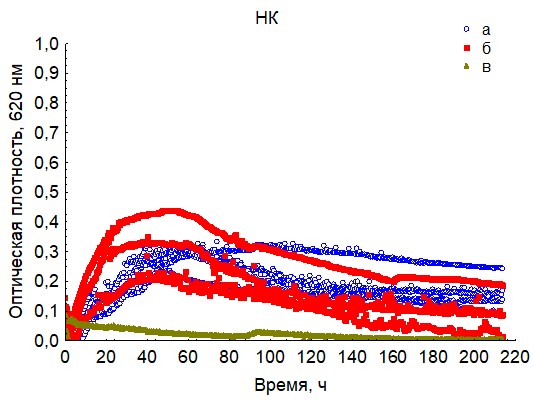
Рис. 15. Кривые периодического роста ассоциаций бактерий на нуклеиновой кислоты (a - корм, б - экскременты, в - контроль на стерильность).
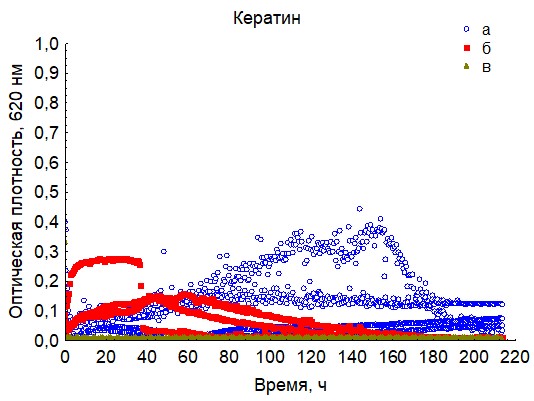
Рис. 16. Кривые периодического роста ассоциаций бактерий на кератине (a - корм, б - экскременты, в - контроль на стерильность).
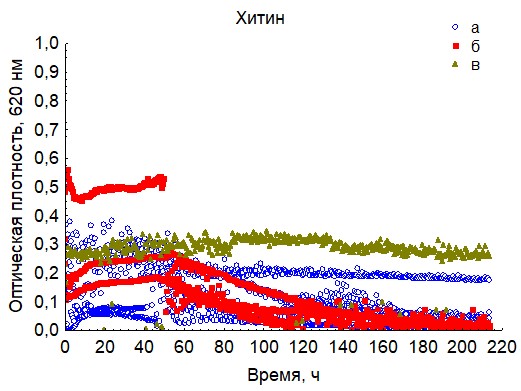
Рис. 17. Кривые периодического роста ассоциаций бактерий на хитине 20 (a - корм, б - экскременты, в - контроль на стерильность).
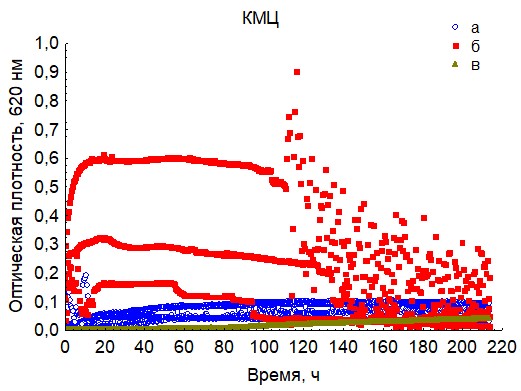
Рис. 18. Кривые периодического роста ассоциаций бактерий на карбоксиметилцеллюлозе (a - корм, б - экскременты, в - контроль на стерильность).
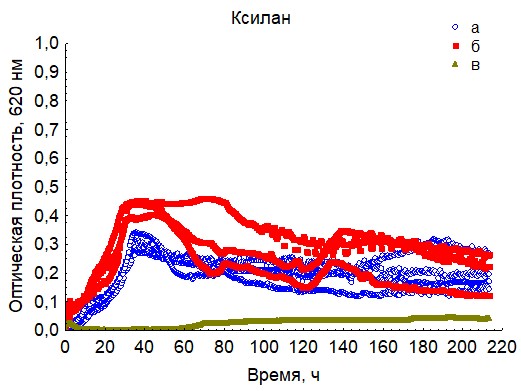
Рис. 19. Кривые периодического роста ассоциаций бактерий на ксилане (a - корм, б - экскременты, в - контроль на стерильность).
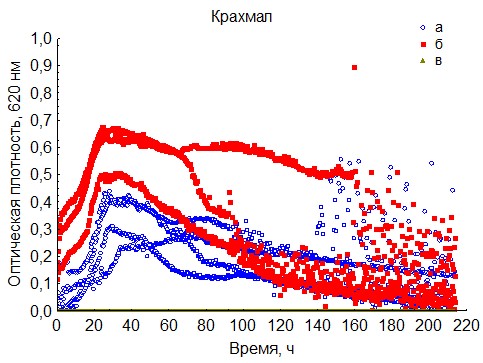
Рис. 20. Кривые периодического роста ассоциаций бактерий на крахмале (a - корм, б - экскременты, в - контроль на стерильность).
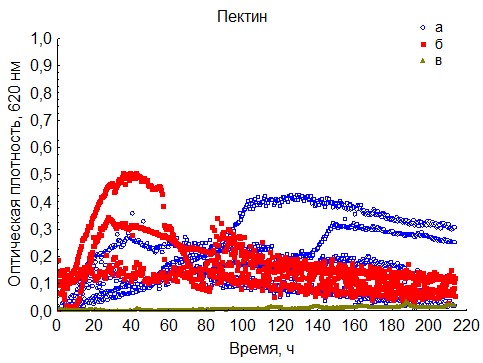
Рис. 20. Кривые периодического роста ассоциаций бактерий на пектиине (a - корм, б - экскременты, в - контроль на стерильность).
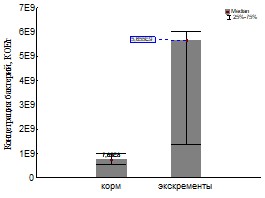
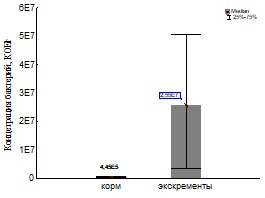
Рис 21 Обилие бактерий по данным посева для кивсяков (слева) и дождевых червей (справа).
Обилие бактерий в исходных корме и экскрементах (Рис.21) исследовалось для определения параметра х0. Однако этот результат имеет и самостоятельное значение. Нами было установлено увеличение обилия культивируемых бактерий при пассаже как дождевых червей так и кивсяков (что согласуется со многими литературными данными (Бызов, 2003)). У дождевых червей увеличение было на два и у кивсяков на один порядок.
Состав ассоциаций, развивающихся на жидких средах с различными биополимерами, определяется путем посева с последующей культурально-морфологической идентификацией бактерий. В табл. 1 представлено количество и соотношение видов бактерий. В табл. 2 приведены индексы Шеннона и Бергера-Паркера характеризующие биоразнообразие для дождевых червей.
Таблица 1. Соотношение видов бактерий формирующих ассоциации на жидких средах с биополимерами.
среда | Корм | копролиты | ||
морфотип № | Доля | морфотип № | Доля | |
крахмал | 3 | 0,72 | 2 | 0,6667 |
4 | 0,26 | 4 | 0,3056 | |
8 | 0,02 | 8 | 0,0278 | |
КМЦ | 13 | 1 | 2 | 0,75 |
9 | 0,25 | |||
пектин | 5 | 1 | 2 | 0,9028 |
10 | 0,0833 | |||
12 | 0,0139 | |||
ксилан | 6 | 0,6667 | 2 | 0,6897 |
11 | 0,3333 | 4 | 0,0689 | |
6 | 0,0345 | |||
8 | 0,2069 | |||
казеин | 1 | 0,6136 | 1 | 0,3443 |
2 | 0,3863 | 2 | 0,1312 | |
3 | 0,5246 | |||
твин 20 | 2 | 0,1064 | 2 | 0,1081 |
8 | 0,2128 | 3 | 0,4595 | |
9 | 0,6809 | 4 | 0,027 | |
8 | 0,3784 | |||
9 | 0,027 | |||
декстран-500 | 2 | 0,8667 | 2 | 0,0919 |
6 | 0,0381 | 4 | 0,4713 | |
7 | 0,381 | 6 | 0,1149 | |
8 | 0,0571 | 8 | 0,0805 | |
10 | 0,2414 |
Таблица 2. Биоразнообразие ассоциаций формирующихся ассоциаций на жидких средах с полимерами.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
