

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
среда | Корм | копролиты | ||
индекс Шеннона | Индекс Бергера-Паркера | индекс Шеннона | Индекс Бергера-Паркера | |
крахмал | 0,9594 | 0,72 | 0,0563 | 0,6667 |
КМЦ | 0 | 1 | 0,81128 | 0,75 |
пектин | 0 | 1 | 0,5176 | 0,9028 |
ксилан | 0,91826 | 0,6667 | 1,27342 | 0,6897 |
казеин | 0,96244 | 0,6136 | 1,40231 | 0,5246 |
твин 20 | 1,19654 | 0,6809 | 1,67436 | 0,4595 |
декстран-500 | 1,12474 | 0,8667 | 1,97424 | 0,4713 |
Результаты табл. 2 были проанализированы методом непараметрической статистики (медиана, квартель) см. рис. 22 и рис. 23. На рис. 24-25 представлены аналогичные графики для кивсяков.
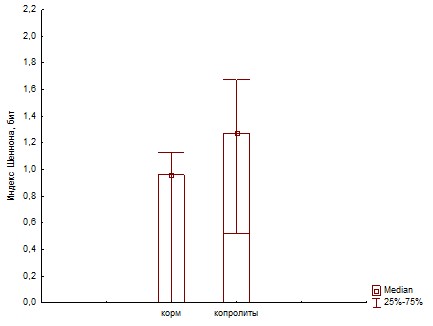
Рис. 22. Сравнение биоразнообразия (индекс Шеннона) для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма и копролитов дождевых червей.
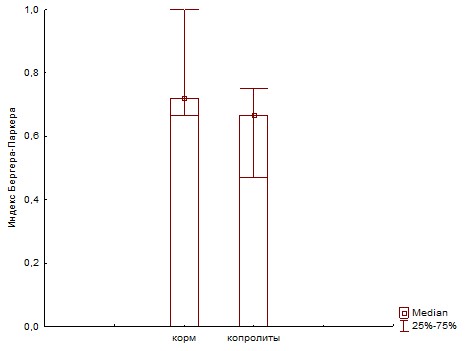
Рис. 23. Сравнение доминирования (индекс Бергера-Паркера) самого обильного вида для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма и копролитов дождевых червей.
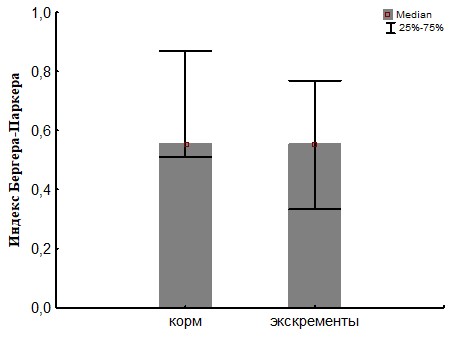
Рис. 24. Сравнение биоразнообразия (индекс Бергера-паркера) для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями подстилки (корма) и экскрементов кивсяков.
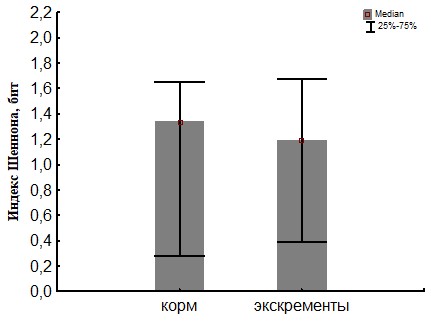
Рис. 25. Сравнение биоразнообразия (индекс Шеннона) для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями подстилки (корма) и экскрементов кивсяков.
При пассаже структурное биоразнообразие бактериального сообщества, оцененное по индексу Шеннона (Рис.22 и 25) снижается за счет увеличения доли доминантного вида, что показывает индекс Бергера-Паркера (Рис.23 и 24).
Интересно охарактеризовать состав ассоциаций на полимерах, по которым мы судим об гидролитическом комплексе непосредственно в корме и экскрементах. Из сообществ, как корма так и экскрементов обоих видов животных на полимерах формируются простые бактериальные ассоциации с индексом Шеннона около 1,3 бит. В них чаще всего доминирует от 2 до 4 морфотипов бактерий. Причем доля выделения новых видов по сравнению с исходной суспензией достигает 77%, а доля общих для корма и экскрементов составляет 41% видов для кивсяков и 54% для дождевых червей. При этом из корма и экскрементов обоих видов животных выделяется на 11 полимерах около полутора десятков морфотипов бактерий.
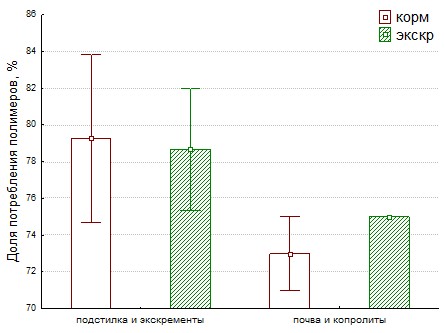
Рис. 25. Доля потребления полимеров в корме и экскрементах бактериями для кивсяков (слева) и дождевых червей (справа).
На основании кривых роста (рис. 3-20) ассоциаций бактерий корма и экскрементов, видны различия между вариантами корма и экскрементов. Исследовался роста на растительных полимерах: ксилан, КМЦ, пектин, крахмал, животных: кератин, микробных: декстран 500, хитин и универсальных: казеин, нуклеиновая кислота и твин 20(аналог жиров). Роста не было на хитине и целлюлозе. На остальных полимерах рост был, однако, не во всех повторностях. Физиологическое разнообразие гидролитического комплекса бактерий для корма и копролитов дождевых червей одинаково (доля от суммарного потребления полимеров 75% и 73% соответственно), как и для кивсяков в подстилке и экскрементах (доля снижается с 80% до 78%) ( Рис.25).
Сравнение физиологического состояния бактерий и преобладающей экологической стратегии роста бактерий в почве (корме) и копролитах.
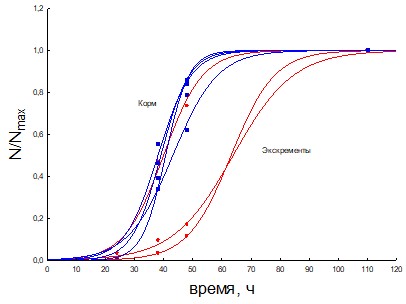
Рис.26. Кривые появления колоний из корм и экскрементов для кивсяков.
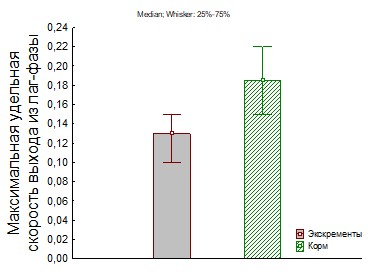

Рис.27. Определение метаболической готовности к росту и интенсивности выхода бактерий из лаг-фазы по динамике появления колоний.
На основание вышеприведенных кривых роста (3-20 рис.) ассоциаций применяя математическую модель роста лаг-фазы и экспоненциальной фазы синтетической хемостатной модели (СХМ) были рассчитаны кинетические параметры мm и г = 100+ln(r0) (показатель метаболической готовности к росту). Результаты определения представлены на рис. 28-35.
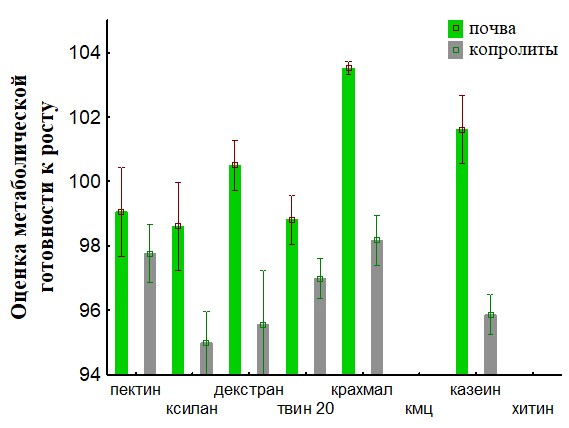
Рис.28. Сравнение метаболической готовности к росту для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма и копролитов для дождевых червей.
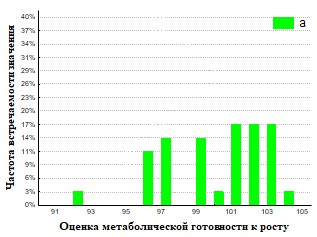
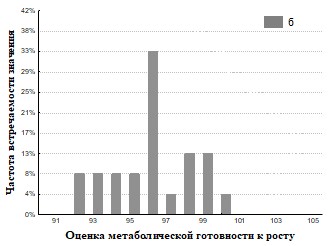
Рис.29. Гистограмма распределения частот встречаемости значений г метаболическая готовность к росту для дождевых червей (а - почва, б - копролиты).
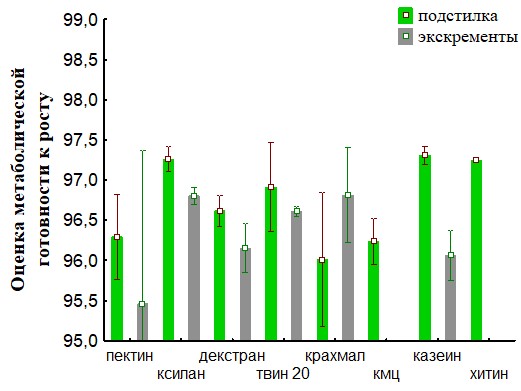
Рис.32. Сравнение метаболической готовности к росту для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма (подстилка) и экскрементов для кивсяков.
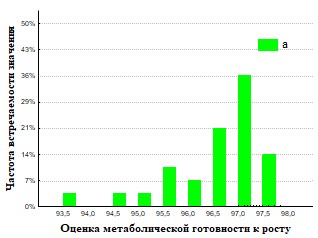
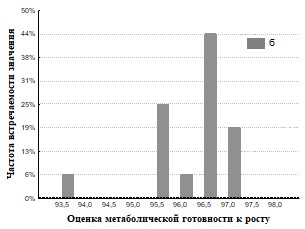
Рис.33. Гистограмма распределения частот встречаемости значений г метаболическая готовность к росту для кивсяков ( а – подстилка, б – экскременты).
На основании полученных данных (Рис.28,29,32,33) представлены результаты определения метаболической готовность к росту. Исследование метаболической готовности микробных ассоциаций расти на питательных средах, позволяет оценить готовность микроорганизмов потреблять полимеры непосредственно в природе. Параметр г (гамма) прямо пропорционален длительности лаг-фаз и чем он больше, тем метаболическая готовность к росту больше. Наша гипотеза состоит в том, что доля метаболически готовых к росту на полимерах ассоциаций (воспринимаемые нами как целое), отражает действительную метаболическую готовность к потреблению полимеров in situ. При пассаже для членов бактериального сообщества наблюдается активизация способности разлагать полимеры в экскрементах кивсяков и инактивация в копролитах дождевых червей.
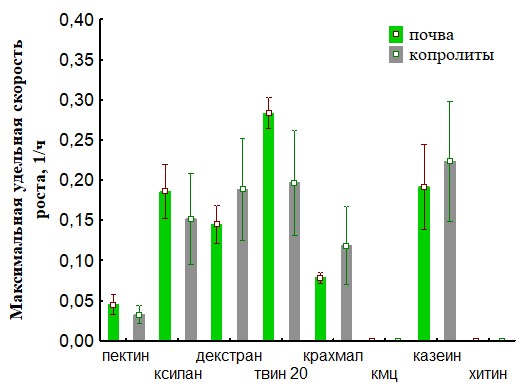
Рис.30 Сравнение максимальной удельной скорости роста для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма и копролитов для дождевых червей.
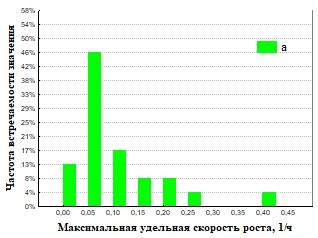
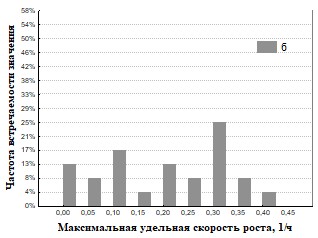
Рис.31 Гистограмма распределения частот встречаемости значений мm максимальная удельная скорость роста для дождевых червей (а – почва, б – копролиты).

Рис.34 Сравнение максимальной удельной скорости роста для ассоциаций развивающихся на биополимерах при инокуляции питательных сред суспензиями корма (подстилка) и экскрементов для кивсяков.
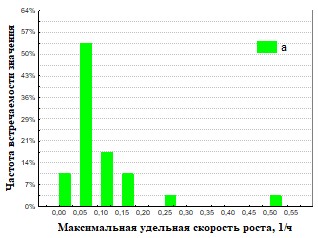

Рис.35 Гистограмма распределения частот встречаемости значений мm максимальная удельная скорость роста для кивсяокв ( а – подстилка, б – экскременты).
На основании полученных данных (Рис.30,31,34,35) представлены результаты определения максимальной удельной скорости роста ассоциаций на полимерах. Наша гипотеза состоит в том, что доля быстрорастущих ассоциаций отражает доли быстрорастущих бактерий (R-стратегов) в сообществе корма и экскрементов. Доля быстро растущих членов бактериального сообщества уменьшается при пассаже у кивсяков и увеличивается у червей. Представление об экологической стратегии сформулировано на основе уравнения логистического роста организмов: 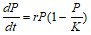 , где r характеризует скорость роста (размножения), а K параметр, характеризующий популяционную ёмкость среды (то есть, максимально возможную численность популяции). Параметры уравнения: r и K послужили обозначению двух основных стратегий роста. Организмы делающие ставку на большие значения показателя r называют r-стратеги, а организмы, которые долгое время поддерживают свою численность на постоянном уровне, называют K-стратегами.. В экологии микроорганизмов существует деление на три группы стратегов: r-стратеги (копиотрофы), K-стратеги (олиготрофы), L-стратеги (гидролитики). В уравнениях описывающих микробный рост аналогом параметра r является мm (максимальная удельная скорость микроорганизмов). L-стратеги характеризуются высокой способностью переживания неблагоприятных условий в покоящемся состоянии (спора) и быстрой активизацией в благоприятных условиях (характеризуется короткой лаг-фазой). Можно сделать вывод, что из копролитов преобладают ассоциации с большим значением мm. Это указывает на большую долю r-стратегов в бактериальном сообществе свежих копролитов по сравнению с почвой. Причина этого в более усиленном разложении органического вещества в кишечнике дождевого червя по сравнению с почвой. Измельчение, перемешивание растительных остатков делает почвенное органическое вещество более доступным для разложения бактериями гидролитиками, которые, выделяя дополнительное количество мономеров, снабжают дождевого червя (хозяина) и копиотрофных бактерий, что стимулирует рост последних.
, где r характеризует скорость роста (размножения), а K параметр, характеризующий популяционную ёмкость среды (то есть, максимально возможную численность популяции). Параметры уравнения: r и K послужили обозначению двух основных стратегий роста. Организмы делающие ставку на большие значения показателя r называют r-стратеги, а организмы, которые долгое время поддерживают свою численность на постоянном уровне, называют K-стратегами.. В экологии микроорганизмов существует деление на три группы стратегов: r-стратеги (копиотрофы), K-стратеги (олиготрофы), L-стратеги (гидролитики). В уравнениях описывающих микробный рост аналогом параметра r является мm (максимальная удельная скорость микроорганизмов). L-стратеги характеризуются высокой способностью переживания неблагоприятных условий в покоящемся состоянии (спора) и быстрой активизацией в благоприятных условиях (характеризуется короткой лаг-фазой). Можно сделать вывод, что из копролитов преобладают ассоциации с большим значением мm. Это указывает на большую долю r-стратегов в бактериальном сообществе свежих копролитов по сравнению с почвой. Причина этого в более усиленном разложении органического вещества в кишечнике дождевого червя по сравнению с почвой. Измельчение, перемешивание растительных остатков делает почвенное органическое вещество более доступным для разложения бактериями гидролитиками, которые, выделяя дополнительное количество мономеров, снабжают дождевого червя (хозяина) и копиотрофных бактерий, что стимулирует рост последних.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
