
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На следующем структурном уровне изображенные на Рис.11-2 суперспирали часто (но не всегда — например, не в тропомиозине) слипаются друг с другом и образуют фибриллы.
Входящие в суперспираль a-спирали обычно параллельны, и перевиты они — в разных белках — по две, три или четыре. У a-спирали, как мы уже знаем, период равен 3.6 остатка на виток. В перевитых спиралях периодичность — 7 остатков на два витка a-спирали, т. е. 3.5 остатка на виток (Рис.11-3, 11-4).
Типичная первичная структура при этом выглядит, как на Рис.11-4.
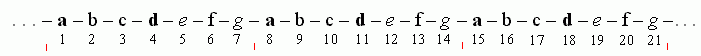 Рис.11-4.
Рис.11-4.
На этом рисунке жирные буквы соответствуют гидрофобным (жирным, слипающимся) аминокислотам, а прочие буквы — аминокислотам гидрофильным. Интересно, что небольшое увеличение гидрофобности остатков e и g превращает двойную перевитую спираль (Рис.11-3а) в тройную (Рис.11-3б), а еще большее — в четверную.
Остановимся чуть подробнее на том, как спирали слипаются между собой. На a-спирали (Рис.11-5, слева) есть несколько спиральных нарезок — "хребтов" из сближенных в пространстве боковых групп. Одни хребты имеют периодичность типа 1 — 4 — 7 — ... , и та их часть, что входит в зону контакта, состоит из пар a1 — d4, a8 — d11, ... (Рис.11-5, в центре). Другие хребты имеют периодичность типа 0 — 4 — 8 — 12 — ... . Та часть этих хребтов, что входит в зону контакта, состоит из пар d4 — a8, d11 — a15, ... (Рис.11-5, справа).
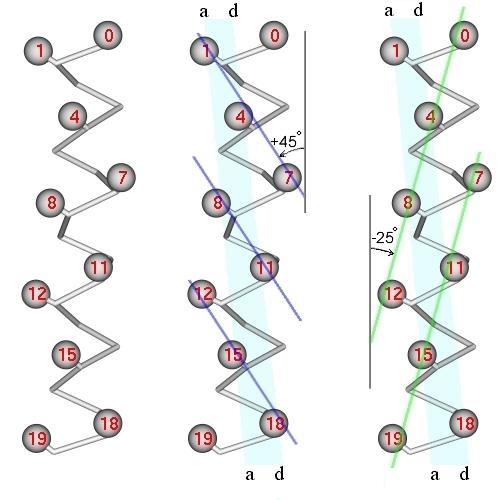
Рис.11-5. a-Спираль (главная цепь и Cb-атомы) и два сорта хребтов (тонкие линии) из сближенных боковых групп на ее поверхности. В центре: хребты типа i, i+3; справа: хребты типа i, i+4. Полосой показана контактная поверхность; ее краями выделены линии остатков a и d. Отмечены типичные углы наклона хребтов i, i+3 и i, i+4 относительно оси спирали (на рисунке углы представляются меньшими, так как типичные хребты проходят через массивные боковые группы, а на рисунке - через центры Cb-атомов).
Первые хребты (если проводить их через выступы, образованные боковыми группами) идут под углом примерно 25о к оси спирали, вторые — под углом примерно 45о (Рис.11-5б и 11-5в). Если, перевернув вокруг вертикальной оси, наложить одну поверхность на другую (Рис.11-6, слева), а затем повернуть на 20о вокруг вертикальной оси, — то хребты типа "1 — 4 — 7" одной спирали окажутся между хребтами типа "0 — 4 — 8" другой, что обеспечит их плотный контакт (Рис.11-6, справа). При этом группы а одной спирали окажутся между группами d другой, а зона контакта образует на поверхности обеих спиралей (пока они не суперспирализованы) слабо скрученную спиральную полосу. А когда эти спирали суперспирализуются (Рис.11-2) и обовьются вокруг общей оси — зона контакта окажется в центре слегка перевитого пучка.
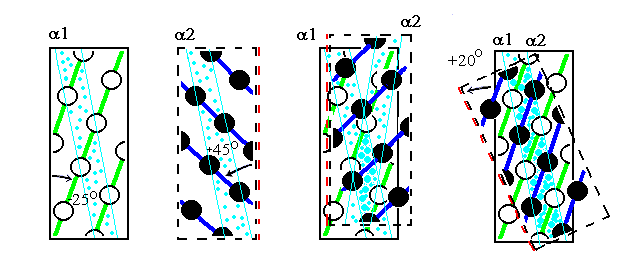
Рис.11-6. Плотная упаковка хребтов боковых групп при контакте спиралей требует разворота на 20о одной спирали относительно другой. Мы смотрим на зону контакта сквозь одну спираль (сквозь перевернутую вдоль оси a2). Остатки "нижней" спирали a1 изображены более светлыми, а верхней (a2) — более темными. Бледно-голубыми линиями показаны линии контактирующих остатков a и d.
Это — не единственный способ плотного контакта спиралей (с другими мы познакомимся, когда будем говорить о глобулярных белках), — но единственный, пригодный для очень длинных спиралей, типичных для фибриллярных белков. Он был предсказан еще Криком в том же 1953 году, когда он — вместе с Уотсоном — предсказал двойную спираль ДНК.
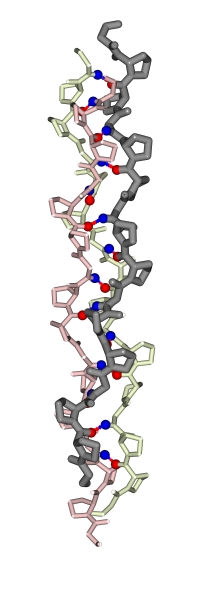
в) Коллаген. Главный опорный белок. Он образуется особой, тройной суперспиралью, сложенной из трех полипептидов (Рис.11-7). При этом внутри каждого полипептида, внутри каждой нити этой тройной спирали водородных связей нет — они есть только между нитями.
Конформация всех остатков в каждой цепи коллагена близка к конформации полипролиновой [точнее — poly(Pro)II] спирали. Это — левая спираль, и период ее равен трем. Соответственно, основной мотив первичной структуры в коллагене выглядит как многократный повтор троек (Gly-Pro-Pro)n, или, точнее, (Gly-нечто-Pro)n, причем Gly в такой тройке необходим для образования водородных связей: у него есть NH-группа (в отличие от Pro), и нет боковой группы, — а любая боковая группа была бы лишней в центре тугой коллагеновой спирали, где сидит глицин.
Интересно, что экзоны, кодирующие коллагеновую цепь, всегда начинаются с глицинов и всегда содержат число кодонов, кратное трем. Я надеюсь, вы помните, что гены эукариот содержат экзоны, кодирующие белки, и интроны, которые выщепляются из матричной РНК (и потому белки не кодируют).
Рис.11-7. Модель тройной суперспирали коллагена для последовательности (глицин — пролин — пролин)n. Каждая цепь выделена своим цветом. Отмечены завязывающие водородные связи Н-атомы NH-групп глицина (синим) и О-атомы СО-групп первого пролина тройки Gly-Pro-Pro (красным). При этом Gly цепи "1" завязывает связь с цепью "2", а Pro — с цепью "3", и т. д. Завиваясь вокруг двух других, каждая цепь коллагена образует правую суперспираль. "Супер" — потому что на более мелком масштабе, на масштабе конформаций отдельных остатков, коллагеновая цепь уже образует спираль типа poly(Pro)II (причем эта "микроспираль" — левая); ее можно проследить по направлению пролиновых колец. |
|
На следующем структурном уровне, коллагеновые суперспирали также слипаются друг с другом и образуют фибриллы коллагена.
Биосинтез коллагена, его последующая модификация и образование зрелой структуры коллагеновой фибриллы хорошо изучены (Рис.11-8). Замечу, что сам по себе коллаген не способен к спонтанной и при этом правильной самоорганизации своей пространственной структуры in vitro — так же, как и фиброин шелка, — и в отличие от глобулярных белков, о которых я буду говорить потом. Для самоорганизации нужен проколлаген, включающий, кроме коллагеновых нитей, глобулярные головки и хвостики. Лишенные головок и хвостиков коллагеновые нити, самоорганизуясь из развернутого состояния in vitro, тоже складываются в тройные спирали — но "неправильные", без присущей нативному коллагену гетерогенности тройной спирали (включающей нити двух сортов), без присущего ему регистра (т. е. с неправильным сдвигом нитей относительно друг друга), и т. д.
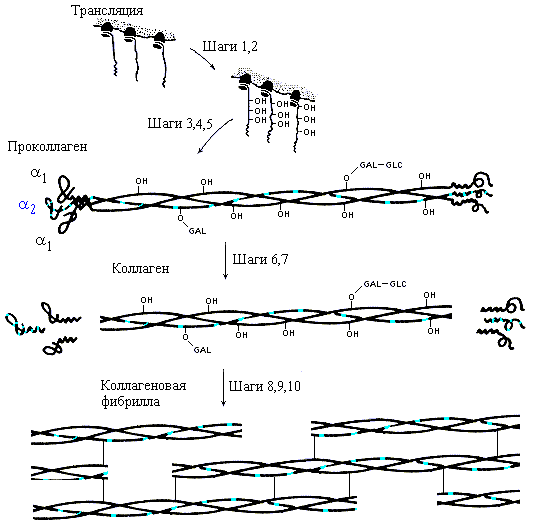
Рис.11-8. Образование коллагена in vivo. Шаг 1. Биосинтез про-a1-цепей и про-a2-цепей (по 1300 остатков в каждой) в пропорции 2:1. Шаг 2. Гидроксилирование некоторых остатков Pro и Lys. Шаг 3. Присоединение сахаров (GLC-GAL) к гидроксилированным остаткам. Шаг 4. Образование тримера и SS связей на его концах. Шаг 5. Образование тройной спирали в середине проколлагена. Шаг 6. Секреция проколлагена во внеклеточное пространство. Шаг 7. Отщепление глобулярных частей. Шаги 8-10. Спонтанное образование фибрилл из тройных суперспиралей, окончательная модификация аминокислотных остатков и образование ковалентных сшивок модифицированных остатков коллагеновых цепей. Картинка взята из [3] и адаптирована.
В заключение, я хочу подчеркнуть, что фибриллярные белки устроены относительно просто в силу периодичности своей первичной и, в силу этого, — также и своей вторичной структуры.
12
Рассмотрим теперь мембранные белки. Они тоже — я имею в виду их трансмембранные части — устроены довольно просто, почти как фибриллярные белки.
Мембраны создают поверхность клетки, а внутри нее, — разные замкнутые объемы — или, как говорят, компартменты. Мембраны состоят из жира (липидов) и белков (Рис.12-1). Особая роль мембранных белков (а они составляют до половины веса мембраны) — обеспечивать транспорт через нее различных веществ, а также сигналов. Мембрана — это, так сказать, "изолятор", а белки — точнее, как мы увидим, каналы в них — "проводники". Эти проводники специфичны, каждый из них пропускает через мембрану только определенные молекулы (или — по-видимому, посредством небольшого изменения конформации белка — сигналы от определенных молекул).

Рис.12-1. Белки в мембране. Серым цветом выделены внемембранные домены. Внутримембранные части белка практически не содержат нерегулярных участков цепи.
Истинно мембранные белки "живут" внутри мембраны — там воды практически нет, так что внутримембранные части таких белков должны состоять (и состоят, как мы увидим) из регулярной вторичной структуры, причем размер этих частей ограничен толщиной мембраны.
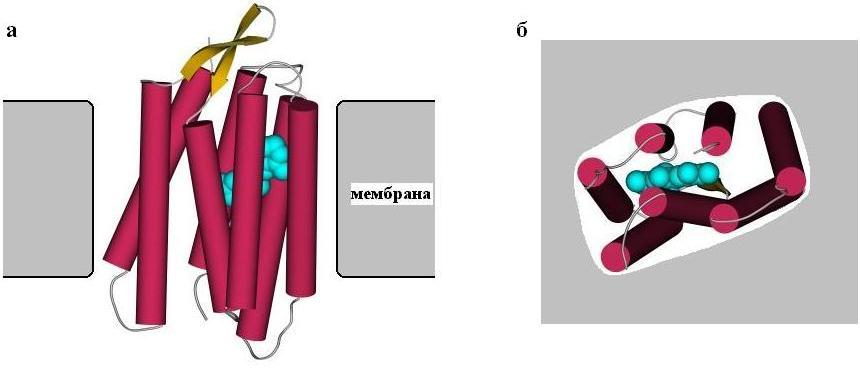
Рис.12-2. Бактериородопсин в мембране (а - вид вдоль мембраны, б - вид на мембрану сверху). Цилиндрами показаны семь спиралей этого белка. Показаны и соединяющие эти спирали петли, а также (голубым цветом) молекула ретинола, прикрепленная внутри бактериородопсина.
Посмотрим, на нескольких примерах, как устроены мембранные белки. Вообще надо сказать, что мы еще знаем мало структур мембранных белков, — их расшифровано меньше десятка, — так как эти белки плохо растворимы в воде (приходится использовать детергенты и т. д.), и их очень трудно кристаллизовать, так как они склонны слипаться неупорядоченно.
Структура бактериородопсина — он проводит протон через мембрану — показана на Рис.12-2; эта структура построена на основе анализа множества электронных микрофотографий очень высокого разрешения, так как получить трехмерные кристаллы бактериородопсина не удалось.
Мы видим, что трансмембранная часть бактериородопсина сложена из семи регулярных a-спиралей, идущих от одного до другого края мембраны и образующих слегка наклоненный к ее плоскости пучок, а одинокая b-шпилька и все нерегулярные участки цепи (соединяющие спирали петли) выходят из мембраны.
Высокая регулярность укладки остова трансмембранной белковой цепи закономерна: в "жирном", почти безводном липидном окружении цена каждой водородной связи очень высока, что заставляет белковую цепь, уж если она попадает в мембрану, принимать структуры с полностью завязанными водородными связями — т. е. либо a-спираль, либо b-структуру (точнее, b-цилиндр, не оставляющий края b-листа открытыми — см. ниже).
Сидящие на a-спиралях бактериородопсина гидрофобные группы обращены "наружу", к (тоже гидрофобным) липидам мембраны. Полярные же группы — их немного — обращены внутрь узкого канала, по которому идет протон. Протонная проводимость осуществляется при содействии прикрепленной внутри пучка спиралей молекулы кофактора — ретинола.
Аналогичные, устроенные как полый пучок спиралей поры могут образовываться в других случаях и из отдельных a-спиральных трансмембранных пептидов.
Посмотрим еще на один трансмембранный белок, порин (Рис.12-3). Он также высоко регулярен и имеет вид широкого цилиндра, сложенного из b-структуры. Обратите внимание, что b-лист здесь образует замкнутый b-цилиндр, что позволяет избежать "незавязанных" водородных связей, типичных для краев плоского b-листа. В этом цилиндре 16 очень длинных b-участков, а диаметр проходящей по его центру довольно широкой поры — около 15![]() . В сторону поры обращены боковые группы полярных остатков, входящих в b-тяжи, а чередующиеся с ними в b-тяжах неполярные остатки обращены своими боковыми группами в мембрану.
. В сторону поры обращены боковые группы полярных остатков, входящих в b-тяжи, а чередующиеся с ними в b-тяжах неполярные остатки обращены своими боковыми группами в мембрану.
Порин служит проводником полярных молекул — кстати, не очень селективным.
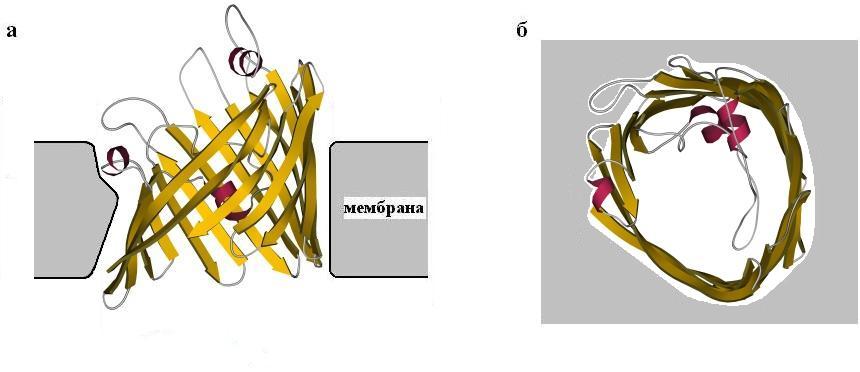
Рис.12-3. Порин. (а) Вид вдоль плоскости мембраны (расположение липидов в ней показано очень схематично). (б) Вид поперек плоскости мембраны.
Селективность проводимости, т. е. специфичность действия мембранных белков во многом связана с тем, что полярные, а тем более заряженные группы "сами по себе" проникают внутрь мембраны с большим трудом.
Вы помните, что свободная энергия заряда q в среде с диэлектрической постоянной e равна +q2/2er, где r — Вандерваальсов радиус заряда. Легко прикинуть, что при q, равном электронному заряду, и r =1.5 ![]() (характерный радиус однозарядного иона), величина +q2 /2er близка к 1.5 ккал/моль при e=80 (т. е. в воде), а при eмембр=3 (т. е. внутри "чистой", сложенной лишь из липида мембраны), величина +q2/2er близка уже к 37 ккал/моль. Итого — повышение свободной энергии на DF = +35 ккал/моль. Вероятность набрать такую свободную энергию есть, по формуле Больцмана, exp(-DF/kT) = exp(-35/0.6) = 10-25. Значит, успешной будет лишь одна из 1025 атак мембраны ионом. А так как "атака" длится не менее 10-13 сек (это — время теплового колебания, как мы помним), то для проникновения иона через чисто липидную мембрану нужно по крайней мере порядка 1012 секунд, или порядка десятка тысяч лет... Все это делает чисто липидную мембрану практически непроницаемой для ионов.
(характерный радиус однозарядного иона), величина +q2 /2er близка к 1.5 ккал/моль при e=80 (т. е. в воде), а при eмембр=3 (т. е. внутри "чистой", сложенной лишь из липида мембраны), величина +q2/2er близка уже к 37 ккал/моль. Итого — повышение свободной энергии на DF = +35 ккал/моль. Вероятность набрать такую свободную энергию есть, по формуле Больцмана, exp(-DF/kT) = exp(-35/0.6) = 10-25. Значит, успешной будет лишь одна из 1025 атак мембраны ионом. А так как "атака" длится не менее 10-13 сек (это — время теплового колебания, как мы помним), то для проникновения иона через чисто липидную мембрану нужно по крайней мере порядка 1012 секунд, или порядка десятка тысяч лет... Все это делает чисто липидную мембрану практически непроницаемой для ионов.
Другое дело, если в мембрану встроен белок, а внутри него образован более или менее широкий, заполненный водой канал, по которому ион может идти, не покидая среду с высокой диэлектрической проницаемостью. Правда, и в канале ион будет чувствовать низкую диэлектрическую проницаемость окружающей канал мембраны, но эффект будет уже много меньше — грубая оценка показывает, что он порядка +q2/(4eмембрR), где R — радиус канала, а eмембр =3. Легко прикинуть, что при этом канал радиусом »2 ![]() в мембране преодолевается ионом за секунду, а »3
в мембране преодолевается ионом за секунду, а »3 ![]() — за долю миллисекунды.
— за долю миллисекунды.
Пропускание иона через канал регулируется наличием в нем центров, способных притянуть данный ион и тем уменьшить барьер, который ему приходится преодолевать.
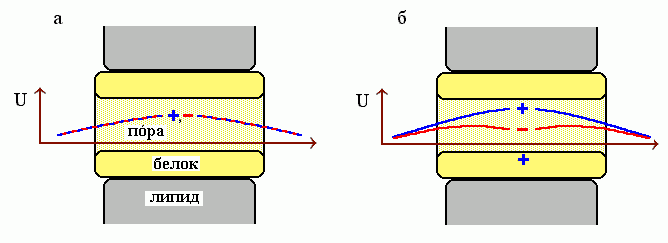
Рис.12-4. Схема трансмембранной поры (мембрана в данном случае расположена вертикально а пора — горизонтально), и электростатическая свободная энергия U для положительно ( .............. + .............. ) и отрицательно ( .............. - ..............) заряженных ионов. (а) На внутренней поверхности поры нет заряда. (б) На внутренней поверхности поры — положительный заряд.
Например, наличие на белке вблизи канала хоть какого-то положительного заряда ускоряет пропускание отрицательно заряженных ионов и резко замедляет пропускание ионов, заряженных положительно (Рис.12-4), а наличие отрицательного заряда — наоборот, ускоряет пропускание положительных ионов и замедляет пропускание отрицательных. Этот эффект служит основой селективности той проводимости, что обеспечивается мембранными белками.
Теперь давайте более подробно рассмотрим фотосинтетический реакционный центр (Рис.12-5). Его задача — проводить выбитые светом электроны на другую сторону мембраны, и тем самым создавать тот электрический потенциал, на котором и работает фотосинтез.
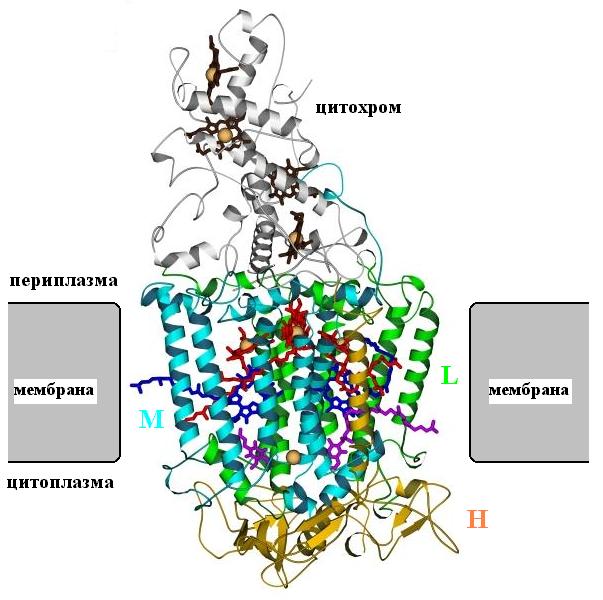
Рис.12-5. Фотосинтетический реакционный центр. Мембрана показана схематично. Трансмембранная субъединица М выделена голубой раскраской, трансмембранная субъединица L — зеленой, субъединица Н (у нее есть одна трансмембранная спиралью) — желтой; цитохром — серый. Обратите внимание, насколко внутримембранные части регулярнее внемембранных! Субъединицы L и М связывают фотосинтетические пигменты — хлорофиллы (красные с желтым ионом магния в центре) и феофитины (темно-синие). И у тех, и у других есть длинные гидрофобные "хвосты", выступающие из белка в мембрану. Субъединицы L, M связывают и два хинона (выделенные фиолетовой окраской). Цитохром, лежащий вне мембраны, включает четыре гема (черно-серые, с оранжевыми ионами железа внутри). Все кофакторы изображены в виде скелетных моделей; см. также Рис.12-6.
Фотосинтетический реакционный центр включает в себя цитохром с четырьмя гемами (это, собственно говоря, — не мембранный белок: он находится вне ее, в периплазматическом пространстве) и три мембранные субъединицы: L, M и H. (от последней, впрочем, в мембране находится только одна a-спираль). Субъединицы L и M очень похожи.
Все трансмембранные части — a-спирали. Они, как всегда, длинны (их длина равна толщине мембраны) и регулярны. Внутри мембраны нет нерегулярных петель. В то же время лежащая вне мембраны цепь значительно менее регулярна и содержит много петель; в общем, она уложена так же, как в "обычных" водорастворимых глобулярных белках, о которых нам еще предстоит говорить.
Обратите внимание на множество небольших циклических молекул, сидящих в белке фотосинтетического реакционного центра: именно по сложенным из них дорожкам, "проводникам" идет электрон (это видно по изменениям электронных спектров этих молекул), а сам полипептид служит только, так сказать, формообразующим изолятором.
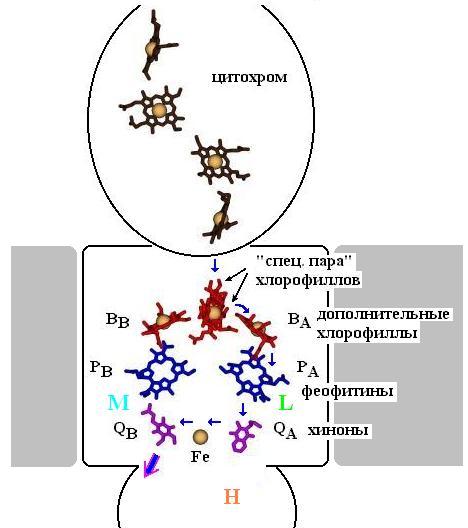
Рис.12-6. Схема расположения фотосинтетических пигментов в фотосинтетическом реакционном центре. Ось псевдосимметрии L и М субъединиц проходит через "специальную пару" хлорофиллов и ион Fe. Длинные "хвосты" пигментов "сбриты", чтобы не загромождать рисунок. Электрон преимущественно идет по правой (на рисунке) ветви пигментов, привязанной к субъединице L. Путь электрона отмечен синими стрелками. Большая фиолетово-синяя стрелка отмечает выход хинона, нагруженного двумя последовательно прибывшими электронами. Левая цепь не используется. Предполагается, что она использовалась в прошлом, а теперь осталась, как аппендикс.
Как же работает фотосинтетический реакционный центр?
Сначала квант света выбивает электрон из сидящей в белке "специальной пары" ("special pair") хлорофиллов (см. схему на Рис.12-6). Этот электрон мгновенно (менее чем за сек) перескакивает на соседний "добавочный" (accessory) хлорофилл, отстоящий на 3![]() от ближайшего хлорофилла "специальной пары", а затем — за ~10-10 сек, через феофитин РА, — на хинон (quinone) QA (именно QA, а не на QB!). Потом — уже за ~10-4 сек — электрон переходит на хинон QВ. Почему электрон идет столь кружным путем на хинон QВ, почему он не регистрируется (по электронным спектрам) на феофитине ВВ — еще неясно.
от ближайшего хлорофилла "специальной пары", а затем — за ~10-10 сек, через феофитин РА, — на хинон (quinone) QA (именно QA, а не на QB!). Потом — уже за ~10-4 сек — электрон переходит на хинон QВ. Почему электрон идет столь кружным путем на хинон QВ, почему он не регистрируется (по электронным спектрам) на феофитине ВВ — еще неясно.
Ушедший со "специальной пары" хлорофиллов электрон возмещается электроном, пришедшим с гема цитохрома. На этом завершается первый полуцикл реакции.
В результате второго аналогичного полуцикла, хинон QB нагружается вторым электроном и уходит с обоими электронами (дважды заряженный, он легче вырывается из мембраны), чтобы принять участие в дальнейшем фотосинтезе.
Итак, фотосинтетический реакционный центр переносит электроны из верхнего (на Рис.12-5, 12-6) в нижний компартмент — против создающейся, в результате, разности потенциалов между компартментами. Коэффициент полезного действия этого реакционного центра — около 50% (т. е. 50% поглощенного света превращается в энергию разделенных зарядов: вполне неплохо!).
Отметим два важных физических момента:
1) И хлорофиллы (Рис.12-7), и феофитины, и хиноны, и другие пигменты включают в себя системы "полуторных" (...-С=C-С=...), т. е. p-электронных связей. Иначе говоря, электронные облака в этих молекулах обобществлены (вследствие Полингова резонанса: ...С=С-... Û ...С-С=...), и электроны бегают по этим молекулам, как по кусочкам металла. Тем самым создается потенциальная яма для электронов, где они "делокализованы", т. е. могут смещаться на расстояния, значительно превышающие диаметр атома (кстати, именно делокализация электронов создает характерную окрашенность пигментов: локализованный в отдельной валентной связи электрон возбуждается коротковолновым ультрафиолетовым светом, а делокализованный электрон — "обычным", более длинноволновым светом).
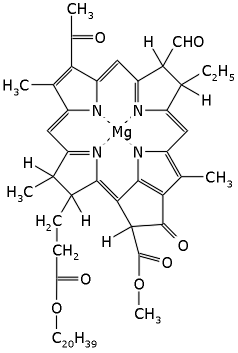
Рис.12-7. Молекула бактериохлорофилла.
2) Передача электрона с одного "кусочка металла" (пигмента) на другой не требует непосредственного контакта этих пигментов. Эта передача осуществляется путем квантового туннельного перехода (см. схему на Рис.12-8).

Рис.12-8. Схема, поясняющая туннельный эффект. Жирная линия показывает профиль потенциальной энергии U электрона. Пунктир — уровень полной (потенциальной + кинетической) энергии электрона. Тонкая линия (со штриховкой под ней) — плотность электронного облака r. Электрон первоначально сидит в левой яме (это его состояние и показано на рисунке), но край его облака (пусть и обладающего здесь очень малой плотностью) достигает правой ямы, так что электрон может со временем весь перетечь туда, если там энергия ниже. При глубине ямы в несколько электрон-вольт (характерной энергии, необходимой для ионизации молекулы) — характерное расстояние, на котором плотность его облака спадает на порядок, — около 1![]() .
.
Суть туннельного перехода (или, как говорят, "подбарьерного" перехода: ведь при этом электрон проходит как бы под энергетическим барьером), — в том, что, по законам квантовой механики, электрон (как и всякая частица, а в особенности — легкая частица) несколько "выступает" за пределы той потенциальной ямы, в которой он находится. "Потенциальной ямой" (областью низкой потенциальной энергии U) в данном случае служит та молекула (хлорофилл, феофитин и т. д.), на которой сидит электрон. Вне "ямы", в которой сидит этот электрон (Рис.12-8), его потенциальная энергия выше, чем его полная (потенциальная + кинетическая) энергия в яме. Если бы не квантовый эффект, этот дефицит энергии не давал бы электрону возможности высовываться из ямы. Квантовый же эффект приводит к тому, что волновая функция электрона (или, попросту, плотность электронного облака) "выплескивается" за потенциальную яму, — хотя величина этой плотности и очень быстро, экспоненциально спадает по мере удаления от "ямы".
Это — тот же квантовый эффект, что не позволяет электрону упасть на ядро: хоть это падение и уменьшило бы потенциальную энергию электрона, его кинетическая энергия возросла бы при этом еще больше. Дело здесь в том, что, при стремлении к нулю расстояния от электрона до ядра (Dx), — потенциальная энергия электростатического взаимодействия электрона с ядром стремится к минус бесконечности как 1/(Dx), а его кинетическая энергия, в силу принципа неопределенности Гайзенберга стремится (при Dx®0) к плюс бесконечности куда быстрее, — как 1/(Dx)2.
Действительно, согласно принципу Гайзенберга, неопределенность в скорости (Dv) и неопределенность в координате (Dx) частицы связаны соотношением mDvDx ~ ![]() , где
, где ![]() — постоянная Планка, а m — масса частицы. Иными словами, абсолютная величина скорости частицы (v) в яме шириной Dx составляет порядка
— постоянная Планка, а m — масса частицы. Иными словами, абсолютная величина скорости частицы (v) в яме шириной Dx составляет порядка ![]() /(mDx) — при полной неопределенности направления движения частицы в данный момент времени. Следовательно, кинетическая энергия частицы, E = mv2/2, есть величина порядка m[
/(mDx) — при полной неопределенности направления движения частицы в данный момент времени. Следовательно, кинетическая энергия частицы, E = mv2/2, есть величина порядка m[![]() /mDx)] 2 = (
/mDx)] 2 = (![]() 2/m)/Dx2.
2/m)/Dx2.
Так же обстоит дело и в случае электрона в потенциальной яме: если бы он ни на йоту не выступал бы за свою яму, его полная энергия была бы выше.
Поэтому электронное облако слегка "выплескивается" за потенциальную яму. При этом плотность его спадает экспоненциально — как и плотность электронного облака атома. Характерное расстояние, на котором плотность этого облака спадает на порядок (в 10 раз) — около 1![]() (это, как мы знаем, — характерный радиус атома).
(это, как мы знаем, — характерный радиус атома).
Значит, плотность электронного облака спадает в 1000 раз на расстоянии в 3 ![]() от "своей ямы". Это означает, что вероятность того, что за одно колебание электрон отойдет от "своей ямы" на 3
от "своей ямы". Это означает, что вероятность того, что за одно колебание электрон отойдет от "своей ямы" на 3 ![]() — порядка 10-3 (а на 5
— порядка 10-3 (а на 5 ![]() , — порядка 10-5, на 10
, — порядка 10-5, на 10 ![]() , — порядка 10-10 и т. д.). Электрон в "яме" (молекуле пигмента) совершает ~1015 колебаний в секунду (в видимом световом диапазоне частот: это хорошо видно по спектрам поглощения таких молекул). Значит, характерное время его перехода в другую "яму" (другую молекулу пигмента), отстоящую на 3
, — порядка 10-10 и т. д.). Электрон в "яме" (молекуле пигмента) совершает ~1015 колебаний в секунду (в видимом световом диапазоне частот: это хорошо видно по спектрам поглощения таких молекул). Значит, характерное время его перехода в другую "яму" (другую молекулу пигмента), отстоящую на 3 ![]() — порядка 10-12 сек; в яму, отстоящую на 5
— порядка 10-12 сек; в яму, отстоящую на 5 ![]() — порядка сек; в яму, отстоящую на 10
— порядка сек; в яму, отстоящую на 10 ![]() — 10-5 сек; а в яму, отстоящую на 15
— 10-5 сек; а в яму, отстоящую на 15 ![]() — порядка 1 сек. Такая зависимость скоростей переходов от расстояний согласуется с тем, что наблюдается в фотосинтетическом реакционном центре.
— порядка 1 сек. Такая зависимость скоростей переходов от расстояний согласуется с тем, что наблюдается в фотосинтетическом реакционном центре.
Обратите внимание на ряд существенных моментов.
Первое. Суммарный переход совершается на расстояние около 40![]() . Такое расстояние нельзя покрыть одним туннельным прыжком (такой прыжок занял бы ~ 1040xсек ~ ~1025 сек ~ 1017 лет — на него не хватило бы всего времени жизни Вселенной). Однако конструкция белка разбивает этот большой прыжок на четыре малых — от одного притягивающего электрон пигмента к другому — и электрон покрывает эти 40
. Такое расстояние нельзя покрыть одним туннельным прыжком (такой прыжок занял бы ~ 1040xсек ~ ~1025 сек ~ 1017 лет — на него не хватило бы всего времени жизни Вселенной). Однако конструкция белка разбивает этот большой прыжок на четыре малых — от одного притягивающего электрон пигмента к другому — и электрон покрывает эти 40![]() за долю миллисекунды.
за долю миллисекунды.
Второе. Для того, чтобы электрон не вернулся бы тотчас со второго пигмента на первый, а пошел бы дальше, на третий и т. д. — его суммарная (потенциальная + кинетическая) энергия должна падать по ходу процесса — или, иными словами, электрон должен на каждом шаге переходить с высокоэнергетической атомной орбиты на низкоэнергетическую. Конструкция фотосинтетического реакционного центра призвана обеспечить такое понижение энергии электрона на пигментах по ходу процесса.
Третье. Совершая туннельный переход, электрон не расходует энергии на преодоление барьера (здесь нет никакого "трения") — однако электрон на каждом шаге идет от более высокоэнергетической атомной орбиты к более низкоэнергетической, его энергия понижается — и она расходуется на то, чтобы сделать туннельный переход "эффективным", т. е. необратимым.
И последнее. Туннельный (или, как говорят, "под-барьерный") переход можно отличить от обычного активационного механизма перехода через энергетический барьер по тому, что на скорость туннельного перехода температура не влияет, а скорость активационного перехода (пропорциональная exp(-DE#/kBT), где DE# — энергия активационного барьера, а Т — температура) резко уменьшается при падении температуры.
Лекция 13
Теперь в фокусе нашего внимания будут находиться глобулярные, а точнее — водорастворимые глобулярные белки. Именно они наиболее исследованы: для сотен — изучена спонтанная самоорганизация, для тысяч — расшифрована атомная трехмерная структура. Поэтому именно их обычно имеют в виду, говоря о "типичных белковых структурах", о "закономерностях, наблюдаемых в строении и самоорганизации белков" и т. д. После этой необходимой оговорки — обратимся к структурам глобулярных белков.
Рентгенструктурные исследования (а позже, — двух - и многомерный ЯМР) позволили установить — за 40 лет интенсивной работы многих десятков лабораторий — атомную структуру порядка 2000 белков (а если считать и мутантные формы, и различные функциональные состояния — то раз в пять больше).
Что можно увидеть в структурах белков — водорастворимых глобулярных белков — с птичьего полета?
Мы видим, что небольшие (из 50 — 150, реже из 200 — 250 остатков) цепи укладываются в компактную глобулу диаметром 25-40![]() (Рис.13-1); и что более крупные белки состоят из нескольких таких субглобул — "доменов" (Рис.13-2). Белковая цепь упаковывается в глобулу так же плотно, как органические молекулы — в кристалл. Это видно и при взгляде на поверхность белка (Рис.13-1а), и на "срезе" белковой глобулы, показанном на Рис.13-1б. Однако, рассматривая белок, нам предстоит рассматривать не плотноупакованные электронные облака (или Вандерваальсовы поверхности) атомов — иначе ничего не будет видно внутри белка — а только очищенные от "мяса" атомов (Рис.13-1в) и даже от боковых групп вообще (Рис.13-1г) скелеты белковых молекул. Поэтому не поддавайтесь часто создаваемому рисунками ощущению "рыхлости" белковых глобул!
(Рис.13-1); и что более крупные белки состоят из нескольких таких субглобул — "доменов" (Рис.13-2). Белковая цепь упаковывается в глобулу так же плотно, как органические молекулы — в кристалл. Это видно и при взгляде на поверхность белка (Рис.13-1а), и на "срезе" белковой глобулы, показанном на Рис.13-1б. Однако, рассматривая белок, нам предстоит рассматривать не плотноупакованные электронные облака (или Вандерваальсовы поверхности) атомов — иначе ничего не будет видно внутри белка — а только очищенные от "мяса" атомов (Рис.13-1в) и даже от боковых групп вообще (Рис.13-1г) скелеты белковых молекул. Поэтому не поддавайтесь часто создаваемому рисунками ощущению "рыхлости" белковых глобул!
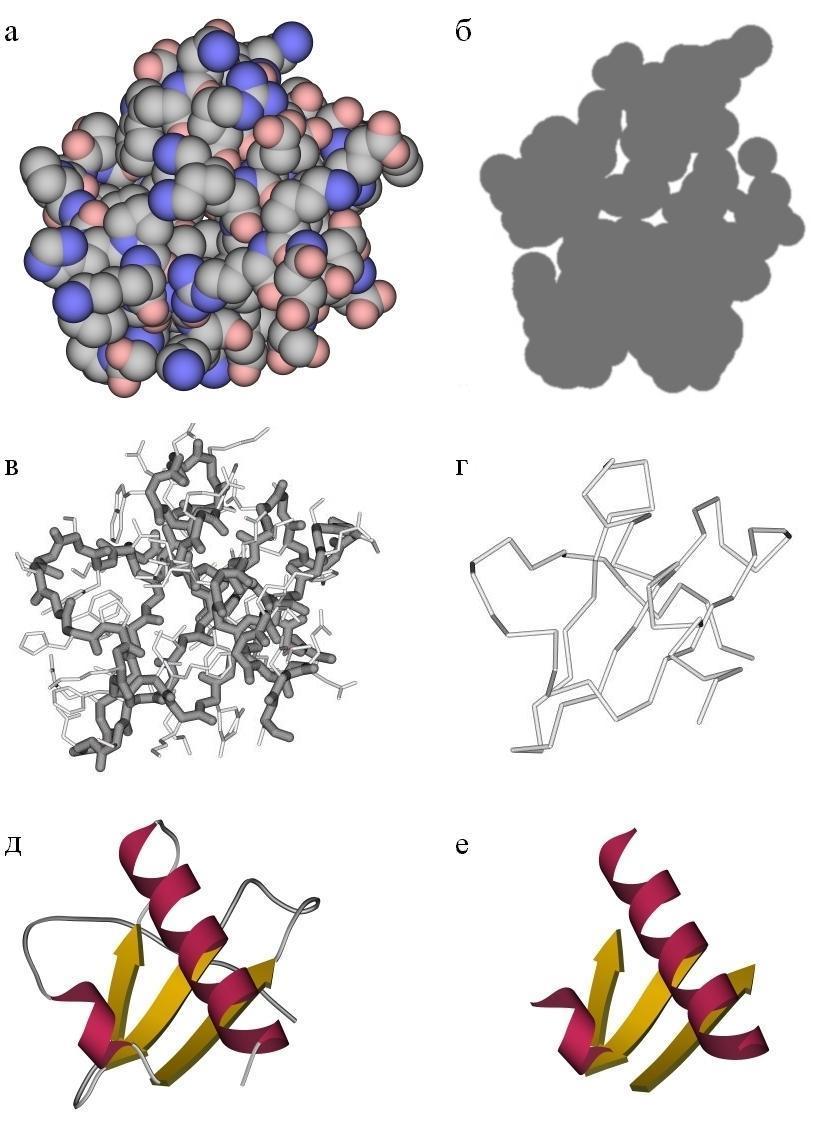
Рис.13-1. Картинки, изображающие строение небольшого белка — a-субъединицы интерлейкина 8 — при разной степени схематизации. (а) Атомная модель (изображены только "тяжелые", не-водородные атомы: синие — азоты, красные — кислороды, серые — углероды); в силу того, что цепь в глобуле упакована плотно, мы видим лишь поверхность белка. (б) Срез атомной модели подчеркивает плотность упаковки. (в) Скелетная модель главной цепи (темная линия) и боковых групп (более светлые отростки). (г) Ход главной цепи. (д) Схема строения белка, на которой выделены вторичные структуры (две a-спирали и один b-лист из трех b-тяжей) в главной цепи белка. (е) Структурный каркас белка, сложенный из вторичных структур. Все рисунки даны в одной и той же проекции и в одинаковом масштабе.
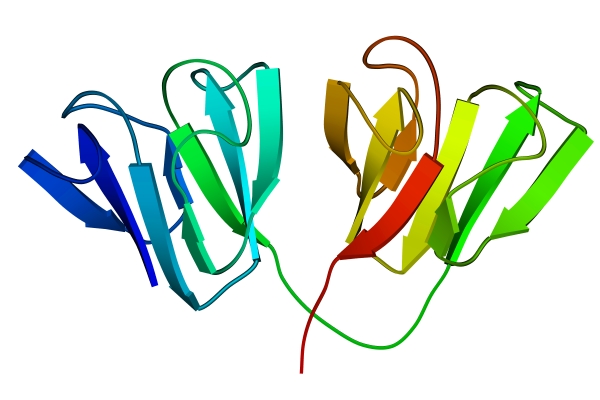
Рис.13-2. Глобулярные домены в g-кристаллине. Цветная кодировка трассирует ход цепи (от синего на N-конце цепи к зеленому в середине и к желтому и красному на С-конце).
Каркас пространственной структуры подавляющего большинства глобул (доменов) сложен из уже изученных нами регулярных вторичных структур — a-спиралей и b-листов (Рис.13-1д), которые стабилизованы регулярными водородными связями в регулярной главной цепи. Кстати, эти вторичные структуры были теоретически предсказаны Полингом, Кори и Брэнсоном еще до расшифровки атомного строения белковых молекул. "Штабель", сложенный из этих структур (Рис.13-1е), определяет основные особенности строения белка.
Каркас из a - и b-структур как бы окружает гидрофобное ядро (или ядра) белка, а нерегулярные петли лежат дальше, на периферии глобулы. Петли практически никогда не входят внутрь белковой глобулы — и неудивительно: их не вовлеченные во вторичную структуру пептидные группы не должны порывать своих водородных связей с водой, это вело бы к нестабильности глобулы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
