
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Как было экспериментально показано, что денатурация небольших белков подчиняется принципу "все-или-ничего" — об этом чуть позже, а пока уместно уточнить, что денатурация типа "все-или-ничего" относится к небольшим глобулярным белкам и отдельным доменам крупных белков, а денатурация больших глобулярных белков, как показывают многочисленные опыты, слагается из плавления входящих в них доменов.
Может ли денатурированный белок ренатурировать и обрести свою нативную структуру? То есть — обратима ли его денатурация?
Да (это известно со времен опытов Анфинсена в 60-х гг.; кстати, за них он получил Нобелевскую премию), — если белок не слишком велик и не подвергался сильной химической модификации после сворачивания in vivo: тогда "мягко" (без разрыва цепи) разрушенная температурой, денатурантом и т. д. — архитектура белка спонтанно восстанавливается при "нормализации" среды. Правда, эффективная ренатурация in vitro нуждается в тщательном подборе экспериментальных условий — иначе ей может воспрепятствовать выпадение из раствора и/или агрегация (межмолекулярная, а для больших белков — видимо, и внутримолекулярная агрегация отдаленных кусков цепи: известно, что способность к ренатурации и ее "выход" обычно падают, а экспериментальные сложности — растут с увеличением размера белка).
Обратимость денатурации белка чрезвычайно важна: она показывает, что нативная структура белка (строго говоря: не модифицированного небольшого белка) — равновесна и стабильна; и эта обратимость позволяет применять термодинамику для описания и изучения такого перехода.
Тот фундаментальный факт, что денатурация белка происходит как переход типа "все или ничего" был показан , работавшим в нашем Институте белка. Он изучал тепловую денатурацию белка, которая обычно сопровождается большим тепловым эффектом, ~1 ккал на моль аминокислотных остатков.
Как доказывается, что плавление белковой молекулы — переход типа "все или ничего"? Для этого недостаточно показать существование пика теплоемкости (или, что есть то же самое, — что энергия белка S-образно зависит от температуры). Все это свидетельствует только о резкости перехода, но ничего не говорит о том, плавится ли белок целиком или по частям. Чтобы доказать, что плавление белка — переход типа "все или ничего", нужно сравнить (1) "эффективную теплоту" перехода, вычисляемую из его ширины, с (2) "калориметрической теплотой" этого перехода, т. е. с количеством тепла, поглощаемым одной молекулой белка в процессе плавления. Совпадение этих двух независимых величин показывает, что молекула плавится как единое целое.
Это и есть "критерий Вант-Гоффа" для наличия перехода типа "все или ничего". Ввиду его важности — рассмотрим этот критерий внимательно.
"Эффективная теплота" перехода, следующая из его ширины, есть количество тепла, поглощенного одной независимой "единицей плавления". Если эффективная теплота перехода меньше калориметрической — "единица плавления" меньше, чем сама молекула, т. е. молекула плавится по частям. Если эффективная теплота перехода больше калориметрической — "единица плавления" больше молекулы, т. е. плавится не одна молекула белка, а какой-то их агрегат.
Как связана эффективная теплота перехода с его шириной? Рассмотрим "единицу плавления", которая может находиться в двух состояниях: "твердом", с энергией Е и энтропией S, и "расплавленном", с энергией Е' и энтропией S'. Пусть, для простоты, E, Е', S и S' не зависят от температуры Т (рассмотрение более общего случая я предоставляю читателю...).
Так как состояний — всего два, то из формулы Больцмана легко видеть, что вероятность пребывания "единицы плавления" в расплавленном состоянии есть
PРАСПЛ = exp[-(E'-TS')/kT] / { exp[-(E-TS)/kT] + exp[-(E'-TS')/kT] } = 1 / { exp[(DE-TDS)/kT] + 1 }, | (16.1) |
где DE=E'-E, DS=S'-S. Вероятность пребывания "единицы плавления" в твердом состоянии есть (для перехода "все-или-ничего") PТВ = 1-PРАСПЛ. Производная dPРАСПЛ/dT показывает, как быстро меняется PРАСПЛ с температурой. Простое вычисление (с использованием уже известной нам формулы d(F/T)/dT = d[(E-TS)/T]/dT = - E/T2) показывает, что
dPРАСПЛ/dT = PРАСПЛ (1-PРАСПЛ) (DE/kT2). | (16.2) |
Пожалуй, надо расписать здесь все преобразования подробнее, чтобы не принимать на веру, а понимать. Обозначим (DE-TDS)/kT через X. Тогда dX/dT = - DE/kT2, PРАСПЛ = 1/(eX+1), PТВ = 1-PРАСПЛ = eX/(eX+1), и
dPРАСПЛ/dT = d[1/(eX+1)]/dT = - [1/(eX+1)2] x [deX/dT] = - [1/(eX+1)2] x eX x [dX/dT] =
= - [1/(eX+1)] x [eX/(eX+1)] x [dX/dT] = PРАСПЛ (1-PРАСПЛ) x (-dX/dT) = PРАСПЛ (1-PРАСПЛ) (DE/kT2)
Точка середины перехода — точка самого быстрого изменения PРАСПЛ, т. е. максимум производной dPРАСПЛ/dT — падает на температуру Т0, где PРАСПЛ=PТВ = 1/2. В этой точке величина PРАСПЛ x (1-PРАСПЛ) проходит через максимум, равный 1/4. Здесь наклон кривой PРАСПЛ(Т) максимален и равен
(dPРАСПЛ/dT)|T=To = (1/4) DE/kT02. | (16.3) |
Графическое определение ширины перехода показано на Рис.16-3, слева. Оно делается путем линейной экстраполяции максимального наклона кривой PРАСПЛ(Т) до пересечения с базовыми прямыми, соответствующими нативному (PРАСПЛ=0) и денатурированному (PРАСПЛ=1) состояниям. В зоне перехода экстраполированная (пунктиром) величина PРАСПЛ |экстрап. меняется от 0 до 1 (то есть DPРАСПЛ |экстрап. = 1), а температура меняется на DT. Значит (dPРАСПЛ/dT)|T=To = DPРАСПЛ |экстрап./DT = 1/DТ, — то есть DТ определяется (см. Рис.16-3) зоной самого крутого подъема величины PРАСПЛ — или, в общем виде, шириной зоны резкого изменения любого экспериментального параметра, определяющегося вероятностью расплавленного состояния (например, шириной зоны резкого изменения спиральности белковой цепи).
В окончательной форме, связь эффективной теплоты плавления "плавящейся единицы" (DE) с шириной (DT) и температурой (T0) перехода имеет вид 1/DT = (1/4) DE/kT02, или
DE = 4kT02/DT. | (16.4) |
Рассчитанная таким образом — только из наблюдаемой формы перехода — теплота DE денатурации "единицы плавления" сравнивается с калориметрической теплотой плавления целого белка (рассчитываемой как DH/N, где DH количество тепла, поглощенного всеми имеющимися в калориметре N молекулами белка). Если
DE = DH/N | (16.5) |
— значит, плавление целого белка является переходом типа "все или ничего". Это и есть "критерий Вант-Гоффа".
Если DE < DH/N (т. е. если ширина перехода DT больше, чем того требует критерий Вант-Гоффа), — то "единица плавления" меньше, чем весь белок, т. е. он плавится по частям. Если DE > DH/N, — то "единица плавления" больше белка, т. е. плавится как целое не одна молекула белка, а какой-то белковый агрегат.
Разрушение структуры белка при повышении температуры — плавление — выглядит естественно; однако существует и "холодовая" денатурация белка при аномальном понижении температуры (Рис.16-4). Правда, для многих белков такая денатурация не наблюдается, так как вода в приборе успевает замерзнуть раньше...
Рис.16-4. Обратимая "холодовая" денатурация белка (апомиоглобина) при аномальном понижении температуры (нижняя кривая; стрелка показывает, что температура в ходе опыта понижается). Верхняя кривая показывает ренатурацию белка (левый пик) при обратном повышении температуры и его дальнейшую денатурацию (правый пик). Кривые относятся к избыточной теплоемкости раствора белка по сравнению с теплоемкостью растворителя; они не отнормированы на количество белка в растворе. Картинка взята из Yu. V.Griko, P. L.Privalov, S. Yu. Venyaminov & V. P.Kutyshenko, J. Mol. Biol. (1988) 202:127-138. |
|
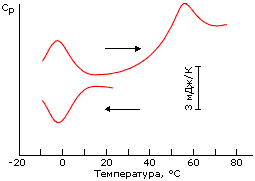
Трудами лаборатории Привалова, где "холодовая" денатурация была впервые обнаружена, было показано также, что она — как и обычная тепловая денатурация — происходит как переход типа "все или ничего".
Причина "холодовой" денатурации кроется в том, что гидрофобные взаимодействия, которые держат белок компактным, сильно растут с температурой — то есть сильно падают с ее понижением. Рост гидрофобных эффектов — и, в частности, их энергии — с температурой проявляется в относительно высокой теплоемкости денатурированного, менее компактного состояния белка (см. DCpd на Рис.16-2). В результате теплота плавления белка сильно растет с температурой — и падает с понижением температуры (Рис.16-2, 16-5а). Падает до того, что может изменить знак при низких температурах (Рис.16-5а)! То есть если обычно более упорядоченная структура (нативный белок) имеет более низкую энергию, чем менее упорядоченная (денатурированный белок) — то при аномально низких температурах (~10оС и ниже) энергия нативного белка может быть выше, чем денатурированного. То же самое, однако, происходит и с энтропией денатурации: она, как и положено энтропии гидрофобного эффекта, тоже падает с падением температуры. В результате стабильность белка (разность свободных энергий его нативной и денатурированных форм) проходит через максимум при "комнатных" или нулевых температурах, — и начинает падать при более значительном охлаждении (Рис.16-5б). При этом иногда стабильность белка падает до того, что при аномально низкой (с точки зрения физиологии: ~0оС и ниже) температуре белок разваливается. Правда, обычно дело до этого не доходит, т. к. вода с белком замерзает, и все процессы крайне замедляются (что позволяет хранить белок на холоду).
Парадоксально, но при холодовой денатурации дело обстоит в точности так, как при кипении воды. Как известно, кипения можно добиться двумя путями — либо подняв температуру, либо понизив внешнее давление. Гидрофобное давление, сжимающее белковую цепь, сильно падает при уменьшении температуры — и белок может "закипеть".
Кстати, действительно "закипеть", — если при повышении температуры белок как бы "плавится", то есть его объем растет не слишком сильно, то при холодовой денатурации белковая цепь совершенно разворачивается и занимаемый ею объем растет многократно.
а | б |
|
|
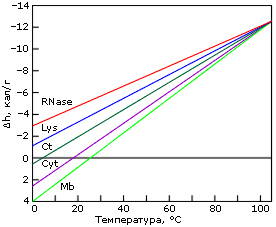
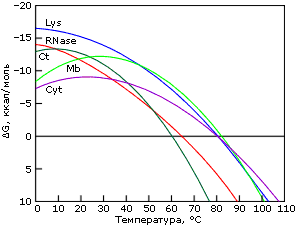
Рис.16-5. Изменение удельной (на грамм белка) разности энергий нативного и денатурированного состояний белка с температурой (а), и изменение свободных энергий в расчете на моль белковых молекул (б). Картинки взяты из P. L.Privalov & N. N.Khechinashvili, J. Mol. Biol. (1974) 86:665-684.
Это естественно подводит нас к вопросу о том, как выглядит денатурированный белок.
Многочисленные термодинамические опыты показывали, что в области денатурированного состояния белковой молекулы в ней нет никаких кооперативных структурных превращений. Поэтому первоначально полагали, что денатурированный белок всегда является очень рыхлым клубком (каковым он, действительно, является в хороших растворителях типа концентрированных растворов мочевины).
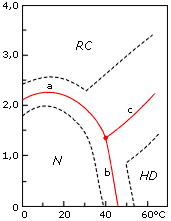
Однако столь же многочисленные структурные исследования денатурированных белков упорно свидетельствовали о каких-то крупных перестройках в рамках денатурированного состояния, о каких-то "промежуточных" структурных состояниях между полностью развернутой (клубковой) и нативной формами молекулы (Рис.16-6).
Вообще, экспериментальные данные о состоянии белковой молекулы после денатурации были очень противоречивы. В одних случаях белки казались полностью "развернутыми" и бесстуктурными, в других — довольно структурированными и/или компактными, а для их полного разворачивания нужны были сильные растворители.
Рис.16-6. Фазовая диаграмма (Танфорд, 1968) конформационных состояний лизоцима при рН 1.7 в растворе гуанидингидрохлорида при различных температурах. N — нативное состояние, RC — клубок ("random coil"), HD — температурно-денатурированное состояние. Жирная линия показывает середину перехода, пунктир — зону перехода (по Танфорду: предположительно, от соотношения 9:1 в пользу одного состояния до 1:9 в пользу другого). Картинка взята из C. Tanford, Adv. Prot. Chem. (1968) 23:121-218. |
|
Разобраться в этой противоречивой картине удалось только, исследуя структуры денатурированных белков целым букетом методов. В этот букет входили и измерения вязкости (дающие объем молекул белка), и спектры КД в дальнем ультрафиолете (отражающие вторичную структуру белка), и спектры КД в ближнем ультрафиолете (отражающие асимметрию, т. е. упорядоченность окружения ароматических боковых групп), — а также и инфракрасная спектроскопия, и ЯМР, и исследование активности белков, и многое другое. Весь этот арсенал и был применен к исследованию денатурации белков группой , куда и я имел честь принадлежать в качестве теоретика.
Оказалось, что, кроме активности белка, есть только две вещи, которые всегда резко меняются при денатурации. Это — (1) упорядоченность окружения боковых групп белка, наблюдаемая как по КД в ближнем ультрафиолете (Рис.16-7а), так и при помощи ЯМР, и (2) фиксированность глобулярной структуры, наблюдаемая — посредством ЯМР — по скорости обмена водородов (Н) полярных групп белка на дейтерий (D) окружающей воды, а также по ускорению протеолиза белковой цепи.
a | б |
|
|
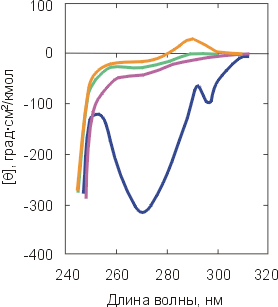
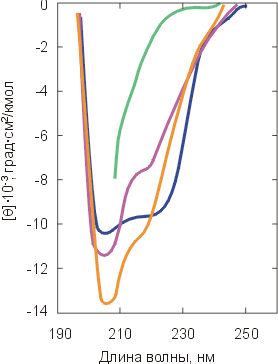
Рис.16-7. Спектры КД в ближнем (а) и дальнем (б) ультрафиолете для нативного a-лактальбумина (![]() ) и для него же, денатурированного теплом (
) и для него же, денатурированного теплом (![]() ), кислотой (
), кислотой (![]() ) и сильным денатурантом (
) и сильным денатурантом (![]() ). Спектры КД в ближнем УФ всегда сильно меняются при денатурации. Спектры КД в дальнем УФ сильно меняются только при сильном разворачивании белка (см. кривую
). Спектры КД в ближнем УФ всегда сильно меняются при денатурации. Спектры КД в дальнем УФ сильно меняются только при сильном разворачивании белка (см. кривую ![]() ). Картинки взяты из D. A.Dolgikh, L. V.Abaturov, I. A.Bolotina, E. V.Brazhnikov, V. E.Bychkova, V. N.Bushuev, R. I.Gilmansin, Yu. O.Lebedev, G. V.Semisontov, E. I.Tiktopulo & O. B.Ptitsyn, Eur. Biophys. J. (1985) 13:109-121.
). Картинки взяты из D. A.Dolgikh, L. V.Abaturov, I. A.Bolotina, E. V.Brazhnikov, V. E.Bychkova, V. N.Bushuev, R. I.Gilmansin, Yu. O.Lebedev, G. V.Semisontov, E. I.Tiktopulo & O. B.Ptitsyn, Eur. Biophys. J. (1985) 13:109-121.
Степень же упорядоченности главной цепи белка (ее вторичная структура), размеры глобулы и даже плотность ее гидрофобного ядра могут как практически сохраняться, так и меняться очень сильно (Рис.16-7б).
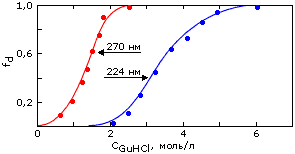
В ходе этих исследований был обнаружен весьма универсальный интермедиат разворачивания (и сворачивания) белков, позже названный "расплавленной глобулой" (Рис.16-8, 16-9). Важнейшие свойства этого интермедиата суммируются Таблицей 16/1.
Рис.16-8. Относительное изменение спектров КД a-лактальбумина в ближнем ( |
|
| Рис.16-9. Схематическая модель нативной и расплавленной глобулы в молекуле белка. Для простоты, на рисунке показаны только две a-спирали, соединенные петлей. Каждая вторичная структура покрыта множеством боковых групп; прошитая водородными связями, вторичная структура стабильна (так что свободная энергия внутримолекулярных водородных связей не меняется), пока проникающий внутрь растворитель не "растворит" всю глобулу. Вода обычно не способна это сделать без добавления денатуранта. Входящие в гидрофобное ядро белка боковые группы заштрихованы. В расплавленной глобуле они обретают свободу движений, выигрывают энтропию, но теряют энергию плотного контакта. Так в глобуле возникают пoры, куда вода ( |
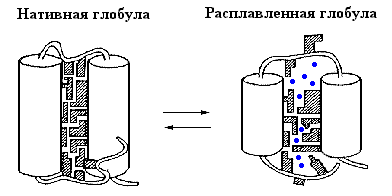
Для очень многих (хотя и не для всех) белков "расплавленная глобула" появляется (путем перехода "все-или-ничего") при умеренном денатурирующем воздействии на нативный белок и исчезает (превращается в неупорядоченный клубок) лишь с добавлением концентрированного денатуранта. "Расплавленная глобула" обычно не плавится при дальнейшем нагреве, но ее разворачивание в клубок сильным растворителем выглядит как кооперативный S-образный переход.
Некоторые нативные белки, однако, разворачиваются прямо в клубок, минуя стадию расплавленной глобулы.
Противоречивость результатов структурных и термодинамических исследований денатурации белков, а также физическая природа нового фазового состояния белков — состояния "расплавленной глобулы" — требовали теоретического осмысления.
Для понимания природы состояния расплавленной глобулы важно, что получается из нативного состояния путем кооперативного температурного плавления — фазового перехода первого рода. Это означает, что оно обладает значительно большей энтальпией и энтропией, чем нативное состояние, то есть что внутримолекулярные взаимодействия в нем резко ослаблены, а подвижность белковой цепи — резко увеличена. Так как большинство внутренних степеней свободы в белке связано с мелкомасштабными флуктуациями структуры, и прежде всего с движениями боковых групп, именно раскрепощение таких флуктуаций может сделать состояние расплавленной глобулы термодинамически выгодным. Однако раскрепощение мелкомасштабных флуктуаций не требует полного разворачивания белковой цепи — достаточно лишь ее небольшого набухания. При этом, однако, в белке резко ослабляется Вандерваальсово притяжение: оно сильно зависит от расстояния и даже небольшого увеличения размеров молекулы достаточно для его значительного ослабления (Рис.16-9).
В общем, здесь все похоже на плавление кристалла: небольшой рост его объема рвет часть Вандерваальсовых взаимодействий в нем и раскрепощает движения молекул.
"Расплавленная глобула" послужила и ключом к пониманию кооперативности денатурации белка. Оказалось, что этот процесс качественно отличается от переходов глобула-клубок в "нормальных" полимерах.
Дело в том, что как эксперименты с гомополимерами (крайне трудные: из-за угрозы агрегации их приходится проводить при предельно низких концентрациях полимера), так и физические теории гомополимеров и статистических гетерополимеров (не "отобранных", как белки, а "нормальных", т. е. "случайных") расходились с тем, что наблюдается в белках.
Они все говорили, что разрушение плотной глобулы в полимерах должно начиниться постепенно, и до самого (или почти до самого) конца идти без резкого скачка плотности молекулы, связанного с переходом одной фазы ("глобула") в другую ("клубок"), что к моменту превращения глобулы в клубок она уже становится очень рыхлой, и что эти две фазы различаются в основном масштабом флуктуаций плотности молекулы (малым — в глобуле, и большим — в клубке), — то есть что этот переход совсем не похож на фазовый переход первого рода — скажем, на испарение. А денатурация белка похожа как раз на плавление или испарение твердого тела.
Несмотря на то, что классическая теория переходов глобула-клубок в гомополимерах не применима к денатурации белков, — я считаю не лишним рассказать о ней — естественно, в самой упрощенной форме. Такой рассказ, с одной стороны, позволит мне позже оттенить то, что существенно именно для белковых, "отобранных" цепей, а не просто для полимеров. С другой стороны — эта теория вполне применима к поведению уже денатурированных белковых молекул. И, наконец, я считаю, что знание базовых физических моделей необходимо для общей культуры.
Я рекомендую вам "поверять слова формулами" при чтении дальнейшего текста. Конечно, проще читать только слова, но они часто допускают неоднозначное толкование; так что, поверяя слова формулами, — вы сами проверите свое понимание.
Итак, я рассмотрю простейшую "полимерную" модель, — много (N>>1) одинаковых и связанных в цепь мономеров ("бусинок"). И я сравню ее с простейшей моделью, используемой со времен Ван дер Ваальса (XIX век!) для описания переходов газ-жидкость — с "облаком мономеров", т. е. с совокупностью из N таких же, но не связанных в цепь мономеров (Рис.16-10).
Облако мономеров | Полимерная цепь | ||
|
|
|
|
|
| ||
|
| ||
T1 < T2 < T3 < T4 | |||
|
|
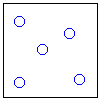
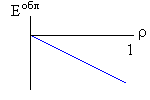
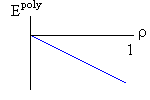
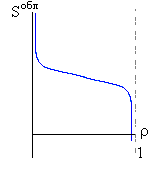
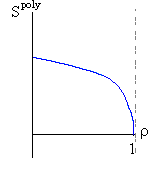
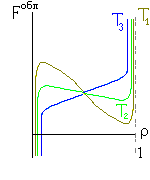
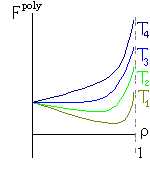
Рис.16-10. Качественное сравнение поведения зависимости энергии DЕ, энтропии DS и свободной энергии DF=DЕ-TDS добавления одного мономера от плотности r для двух систем: "облако мономеров" и "полимерная цепь". Графики для энергий приведены для притягивающихся (e<0) мономеров. Зависимости DF от r показаны для разных температур Т. На дополнительных графиках (внизу) показана качественная зависимость плотности r обеих систем от энергии взаимодействия мономеров, выраженной в единицах kT; графики относятся к невысокому внешнему давлению [при очень высоком, "надкритическом" давлении разреженных (с r<0.5) состояний не существует].
Предварительное замечание. Ниже я буду считать, что мономеры притягиваются (иначе слишком скучно: плотная фаза не образуется, и все), и что интенсивность этого притяжения не зависит от температуры, т. е. что оно имеет чисто энергетическую природу. Последнее часто неверно, так как мономеры плавают в растворителе (вспомните о гидрофобном эффекте: он растет с температурой!), но так легче найти и рассмотреть все основные сценарии возможных фазовых превращений. А потом уже можно усложнить задачу — учесть зависимость "энергии" притяжения от температуры — и указать, когда, при каких температурах, какой из этих сценариев реализуется.
Сначала я хочу дать качественную картину и объяснить, почему "облако" может претерпевать скачок из плотной в разреженную фазу, а полимер — не может, т. е. почему он должен сгущаться постепенно.
Чем жизнь мономера в полимере отличается от его жизни в облаке? Тем, что в полимере он не может далеко отойти от своего соседа по цепи; а в облаке он может отлететь от любого другого мономера куда угодно — пока не наткнется на стенку сосуда.
Почему из мономеров образуется плотная фаза — будь то жидкость (если мономеры не связаны цепью) или глобула (если связаны)? Потому, что мономеры притягиваются друг к другу, и — при низкой температуре, когда роль энтропии падает (вспомните: в свободной энергии F=E-TS энергия E сравнивается с энтропией умноженной на температуру) — энергия притяжения побьет любое желание мономеров рассредоточится по пространству и выиграть энтропию. Побьет всегда — кроме одного случая: когда такое рассредоточение приведет к практически бесконечно большому энтропийному выигрышу. А последнее возможно тогда, когда облако мономеров помещено в очень большой объем. И вот здесь-то мономерам этого облака приходиться выбирать между двумя фазами: либо быть каплей (низкая энергия, но и малая энтропия), либо быть разреженным газом (ничтожная энергия, — но зато очень большая энтропия: летай, где угодно!). А вот промежуточные состояния — с не очень низкой энергией и с невысокой энтропией — будут иметь более высокую свободную энергию, и потому не будут стабильны. Так что стабильной — при низкой температуре — будет либо очень плотная фаза облака, либо очень разреженная, — но не нечто рыхлое. Иначе говоря, — при изменении внешних условий (скажем, при повышении температуры) в "облаке мономеров" может наблюдаться резкое испарение плотной фазы.
Так обстоит дело в "облаке". А в полимере? Не так, — потому что в самой разреженной глобуле мономер никак не может далеко отойти от своего соседа по цепи, — то есть его энтропия в самом разреженном клубке никогда не превзойдет какого-то предела и не сможет (при низкой температуре) конкурировать с энергией, выигрываемой при сжатии глобулы. Так что полимеру не из чего выбирать: при низкой температуре быть ему только сжатым в глобулу, и, значит, — в отличие от "облака" — он не может претерпеть скачок из плотной в разреженную фазу. То есть при изменении внешних условий (скажем, температуры) возможно только постепенное разрыхление (или уплотнение) полимерной глобулы, но никак не ее резкое "испарение".
Теперь повторим то же самое на языке формул.
Сравним "облако" из N мономеров, помещенных в объем V, с глобулой того же объема, образованной цепью из также N мономеров. Считая, что мономеры и "облака", и полимера более или менее равномерно распределены по объему — добавим к каждой из этих систем еще один мономер и посмотрим, как зависит свободная энергия этого мономера (или, как говорят, его химический потенциал) от плотности системы.
Рассмотрим сначала энергию мономера в каждой системе — точнее, ту ее часть, что зависит от плотности, т. е. от взаимодействия ковалентно не связанных звеньев.
В обоих случаях энергия системы (и полимера, и "облака") меняется — при добавлении мономера — на одну и ту же величину. Она меняется на величину энергии взаимодействий добавленного мономера, DE=er, где r=N w/V — удельная плотность системы, т. е. доля ее объема V, занятая N мономерами (с объемом w у каждого), а e — энергия всех взаимодействий мономера в предельно сжатой системе (при r=1). Мы будем считать, что e<0, т. е. что мономеры притягиваются.
Итак, энергии меняются одинаково — а вот энтропии полимера и облака мономеров меняются по-разному.
В "облаке" одному мономеру предоставлен объем V/N. Точнее, — так как прочие мономеры занимают объем Nw , — свободный объем для мономера в "облаке" составляет
V1обл = (V - N w)/N = (V/N)(1 - r) = (w/r)(1 - r). | (16.6) |
Обратите внимание, что этот объем неограниченно велик при стремлении плотности r к 0.
Не то в полимере. Здесь "новому" мономеру доступен лишь ограниченный объем
V1поли = W(1-r) , | (16.7) |
причем объем W ограничен связью мономера с предшествующим звеном цепи, а фактор (1-r), как и раньше, учитывает, что прочие звенья занимают часть r всего объема.
Результат: изменение энтропии при добавлении одного мономера к "облаку" составляет
DSобл = k ln[V1обл]= k ln[(w/r)(1-r)], | (16.8) |
а к цепи -

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
