
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
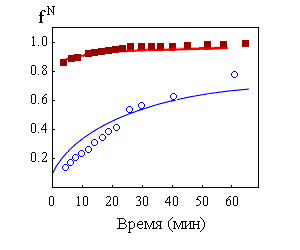
Рис.17-3. Кинетика восстановления "степени нативности" fN в процессе ренатурации карбоангидразы В. Переход белка из полностью развернутого состояния (существовавшего при 5,45 М гуанидингидрохлорида) в нативное (при 0,97 М гуанидингидрохлорида) наблюдается по различным параметрам: приведенной вязкости ![]() , эллиптичности при 222 нм
, эллиптичности при 222 нм ![]() и 270 нм ( _______ ) и энзиматической активности (о о о). Картинка взята из D. A.Dolgikh, A. P.Kolomiets, I. A.Bolotina & O. B.Ptitsyn, FEBS Letters (1984) 164:88-92.
и 270 нм ( _______ ) и энзиматической активности (о о о). Картинка взята из D. A.Dolgikh, A. P.Kolomiets, I. A.Bolotina & O. B.Ptitsyn, FEBS Letters (1984) 164:88-92.
Расплавленная глобула оказывается — в физиологических условиях — ранним промежуточным состоянием на стартующем из клубка пути самоорганизации многих глобулярных белков in vitro. Для образования такого интермедиата достаточно миллисекунд, полное же восстановление свойств нативного белка из 100-300 аминокислотных остатков требует от секунд (для одних белков) до десятков минут (для других).
Необходимо подчеркнуть, что самый медленный (rate-limiting) шаг самоорганизации приходится не на раннюю стадию, не на образование расплавленной глобулы, а на образование из нее плотно упакованной, нативной глобулы (Рис.17-3).
Все описанные выше работы велись, как теперь говорят, в рамках "химической логики", императив которой — "ищи и выделяй промежуточные состояния!". В химии это обычно позволяет понять, как протекает реакция. Но при изучении самоорганизации белков эта логика не сработала: интермедиаты были определены, но ключевые вопросы остались без ответа.
Тогда обратились к маленьким (из 50-100 остатков) белкам, сворачивающимся in vitro без наблюдаемых, "накапливающихся" в эксперименте интермедиатов, и начали исследовать их более интенсивно. И первая неожиданность, с которой здесь столкнулись, — оказалось, что эти белки порой сворачиваются очень быстро — много быстрее тех, наличие интермедиатов в сворачивании которых должно было, казалось бы, ускорить и облегчить их правильное сворачивание!
Обычно белки "однодоменного" размера сворачивались за секунды или десятки секунд, но некоторые из этих белков (правда, это были маленькие белки — до 100 остатков длиной) сворачивались — полностью, до нативной структуры, без наблюдаемых накапливающихся промежуточных структур — за миллисекунды (Рис.17-4). Это регистрировалось по одинаковой скорости восстановления вторичной структуры, упаковки боковых групп, флюоресценции триптофанов (зависящей от их погруженности в глобулу), обмениваемости водородов в белке и т. д.

Рис.17-4. Ренатурация ACBP (белка, связывающего ацил-коэнзим А), регистрируемая по восстановлению эллиптичности при 225 нм (а) и 286 нм (б). Тонкая, ломаная линия — экспериментальный сигнал, жирная кривая — результат его сглаживания, с учетом погрешности опыта. Картинки взяты из Kragelund B. B., Robinson C. V., Knudsen J., Dobson C. M. & Poulsen F. M. Biochemistry (1995) 34: .
Однако, — что в таких экспериментах можно исследовать, чтобы пролить свет на природу процесса самоорганизации? Ведь интермедиатов, которые можно выделить и изучать, тут нет?!
Ответ: здесь можно изучать переходное состояние — состояние, играющее ключевую роль в кинетике процесса.
Напомню, что кинетика перехода между двумя непосредственно наблюдаемыми состояниями (А и В на Рис.17-5) успешно описывается теорией переходных состояний — которая в химии обычно называется теорией активированных комплексов. Суть ее сводится к тому, что скорость процесса лимитируется переходным состоянием #, т. е. наименее (по термодинамике) вероятным (и потому не накапливающимся и непосредственно не наблюдаемым), но необходимым промежуточным состоянием пути из А в В, — или, иными словами, что кинетика переходов А®В (и В®А) определяется высотой максимума свободной энергии на пути, соединяющем А и В.
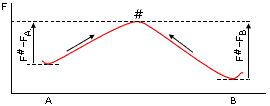
Рис.17-5. Преодоление свободно-энергетического барьера # при переходе из состояния "А" в состояние "В" (стрелка слева) и из "В" в "А" (стрелка справа). FA, FВ и F# — свободные энергии состояний А, В и "переходного" (барьерного, имеющего максимальную свободную энергию на пути процесса) состояния #.
При этом скорости перехода из А в В и из В в А суть
kА®В = k0 exp[-(F#-FA)/RT] kВ®А = k0 exp[-(F#-FВ)/RT]. | (17.1) |
Здесь FA, FВ и F# — свободные энергии состояний А, В и "переходного" # (Рис.17-5), а k0 — быстрота "элементарного шага" процесса, за которую часто принимают частоту теплового колебания RT/h (где h — постоянная Планка; при "обычных" температурах T~300oK, RT/h ~ 1013 сек-1), — с такой частотой, под воздействием тепловых колебаний, молекула предпринимает попытки преодолеть активационный барьер #.
Уравнения (17.1) применимы только к процессам, протекающим за время, много-много большее, чем время элементарного шага. И именно с такими процессами мы имеем дело при сворачивании белков.
Напомню, откуда следуют соотношения (17.1). Рассмотрим процесс А « # ® В. Так как "проток" молекул через барьер # идет медленно по сравнению со скоростью элементарного шага, то состояние # находится почти в термодинамическом равновесии с состоянием А. Иными словами, если в исходном состоянии А находится n молекул, то, по распределению Больцмана, в барьерном состоянии # находится n# = n exp[-(F#-FA)/RT] молекул. За время "элементарного шага" они скатятся с барьера # — половина в сторону А, половина в сторону В [а на их место придет примерно столько же (n#) молекул со стороны А — так что населенность # практически не изменится, будет квазистационарной]. Иначе говоря, для перехода половины всех n молекул через барьер в сторону В потребуется время, в (n/2)/(n#/2) = exp[+(F#-FA)/RT] раз превосходящее время элементарного шага. А поскольку время элементарного шага t0º1/k0, то характерная скорость реакции А « # ® В есть kА®В = k0 exp[-(F#-FA)/RT], а ее характерное время, — время, за которое около половины всех молекул перейдет из А в В, — есть
tА®В = 1/kА®В = (1/k0) exp[+(F#-FA)/RT]. | (17.2) |
Это же объяснение распространяется, естественно, и на kВ®А.
Важно, что температурная зависимость скорости реакции дает возможность судить об энергии переходного состояния. Для этого, согласно Аррениусу, вычисляют производную логарифма скорости реакции по обратной температуре (1/T):
d[ln(kА®В)] / d(1/T) = d[ln(k0) - (F#-FA)/RT]/(-T-2dT) ® - (E#-EA)/R . | (17.3) |
Здесь я воспользовался уже известной нам формулой d(F/T)/dT = - E/T 2 и пренебрег очень слабой зависимостью скорости элементарного шага от температуры. Последнее допустимо даже в простых химических реакциях, и тем более допустимо в белках, где энергии E# и EA — велики, так как определяются взаимодействиями множества частиц.
Рисунок 17-6 показывает изменение скорости сворачивания и разворачивания лизоцима при его тепловой ре - и денатурации.
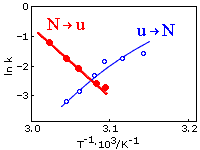
Рис.17-6. Аррениусовы графики для зависимости скорости тепловой денатурации и ренатурации лизоцима от обратной температуры (Т-1); графики взяты из статьи S. Segava & S. Sugihara, Biochemistry (1984) 23:. Константы скорости (k) измеряются в сек-1. Ренатурация, скорость ku®N (эксперимент: точки о, и тонкая интерполяционная кривая); денатурация, скорость kN®u (эксперимент: точки ·, и жирная интерполяционная кривая). Середине плавления отвечает та температура, при которой ku®N = kN®u, т. е. где кривые пересекаются (около 1000/3.08 = 325оК). При меньших температурах (т. е. при больших Т-1 — справа от точки пересечения) превалирует сворачивание, при больших температурах (т. е. при меньших Т-1, слева от точки пересечения) — идет разворачивание.
Он показывает, что в самой середине перехода [там, где скорость сворачивания (ku®N) равна скорости разворачивания (kN®u), так что кривые для ln(ku®N) и ln(kN®u) пересекаются] — скорости обоих процессов близки к е-2.5 » 0.1 сек-1 (т. е. что здесь времена протекания и де-, и ренатурации — порядка 10 сек).
Кроме того, эта картинка показывает, что денатурация ускоряется по мере углубления в "область денатурации", а ренатурация ускоряется по мере углубления в "область ренатурации". И здесь содержится одна очень любопытная вещь.
Она состоит в том, что скорость тепловой денатурации kN®u падает с Т-1 (т. е. что она растет с температурой Т, что обычно для физико-химических реакций), а скорость сворачивания ku®N — наоборот, растет с Т-1 (т. е. она падает с температурой, что необычно для физико-химических реакций). По формуле (17.3) это означает, что E#-EN > 0, а E#-Eu < 0. Иначе говоря, Eu > E# > EN, т. е. энергия барьера, E#, лежит выше энергии нативного состояния, EN, но ниже энергии развернутого состояния, Eu (последнее необычно для химических реакций, где барьер по энергии выше чем и начальное, и конечное состояние реакции). Дальнейший анализ этого графика показывает, что такое же соотношение, Su > S# > SN, справедливо и для энтропий развернутого, переходного и нативного состояний при сворачивании белка. Это значит, что барьер между нативным и развернутым состояниями белка выглядит "обычным", энергетическим барьером, — если глядеть на него со стороны нативного состояния, и что он выглядит необычным, энтропийным барьером, — если глядеть на него со стороны развернутого состояния.
Впрочем, этого можно было ожидать, исходя из сделанного на прошлой лекции анализа свободно-энергетического барьера и причин фазового, типа "все-или-ничего" перехода между нативным и денатурированным состоянием белка (см. Рис.17-7, который воспроизводит уже известную вам схему изменения энергии, энтропии и свободной энергии по мере расширения глобулы).
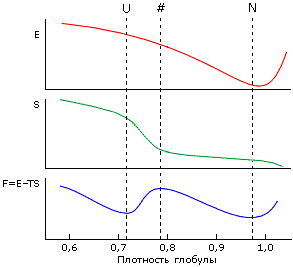
Рис.17-7. Изменение энергии E, энтропии S и свободной энергии F=E-TS с изменением плотности глобулы. D — денатурированное состояние (в данном случае — расплавленная глобула, так как плотность его велика), N — нативная глобула, # — "барьер", максимум свободной энергии на пути равномерного расширения глобулы (от N к D).
Обратимся еще раз к уравнениям (17.1). Они показывают, что отношение скоростей прямой и обратной реакций, kА®В/kВ®А, есть просто константа равновесия между финальным (В) и начальным (А) состояниями
КВ:А = kА®В/kВ®А = exp[-(FВ-FA)/RT] . | (17.4) |
Величина константы равновесия КВ:А есть nB¥/nА¥, она показывает то соотношение между финальным числом nB¥ молекул в состоянии В и их финальным числом nA¥ в состоянии А, к которому, в конечном итоге (при стремящемся к бесконечности времени наблюдения), приходит процесс при данных условиях (температуре и т. д.).
С какой скоростью система приходит к этому равновесному состоянию? Для ответа решим соответствующее дифференциальное уравнение:
dnA/dt = - kА®В nA + kB®A nB | (17.5) |
(второе уравнение, для dnB/dt, писать не надо, т. к. nA+ nB º n0, где n0 — полное число молекул, т. е. dnВ/dt º -dnА/dt). Ответ — позвольте мне написать его без выкладок, а вам я рекомендую сделать их самостоятельно — таков:
nA(t) = [nA(0) — nА¥] exp[-(kА®В + kB®A)t] + nА¥ . | (17.6) |
Здесь nA(t) — число молекул в состоянии А в момент времени t от начала процесса, а nА¥ = n0.[kВ®А / (kА®В + kВ®А)] — конечное, равновесное число молекул в состоянии А.
Значит, видимая (apparent) скорость приближения к равновесию есть
kapp = kА®В + kВ®А, — | (17.7) |
она равна сумме скоростей прямой и обратной реакций. Обратите внимание, что эта скорость зависит только от условий, в которых протекает релаксация, и не зависит от исходного распределения белковых молекул между нативным и денатурированным состояниями [т. е. от исходной доли nA(0)].
В этой суммарной скорости kapp доминирует более быстрая реакция.
Если условия (температура и т. д.) отвечают большей устойчивости нативного состояния, — то в kapp = ku®N + kN®u доминирует ku®N, скорость сворачивания. Если более устойчиво денатурированное состояние, — то в ku®N + kN®u доминирует kN®u, скорость разворачивания.
Большим преимуществом измерения скорости прихода к равновесию, kapp, является то, что она всегда может быть измерена (в отличие от ku®N и kN®u в отдельности), — как тогда, когда более стабильно нативное состояние, так и тогда, когда более стабильно денатурированное.
При измерении kapp строятся так называемые "шевронные графики" (Рис.17-8). Они так называются из-за своей характерной V-образной формы, напоминающей шевроны на военных кителях. Один взгляд на этот график показывает, что скорости сворачивания (ku®N) и разворачивания (kN®u) зависят от концентрации денатуранта противоположным образом. С аналогичной ситуацией мы уже сталкивались, когда изучали температурную зависимость скоростей сворачивания и разворачивания белка. Как и тогда, наблюдаемая здесь противоположность наклонов означает "промежуточность" свойств переходного состояния, их среднее положение между свойствами нативного и развернутого белка.
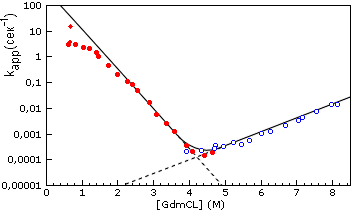
Рис.17-8. "Шевронный график" зависимости kapp = ku®N + kN®u, видимой характерной скорости приближения к равновесию между нативной и развернутой формами белка (лизоцима куриного яйца), от концентрации гуанидингидрохлорида. Заполненные кружки получены при разбавлении раствора GdmCl, в котором изначально находился денатурированный белок, т. е. при полной или частичной ренатурации белка. При этом kapp » ku®N. Пустые кружки получены при добавлении GdmCl к изначально нативному белку, т. е. при полной или частичной его денатурации. При этом kapp » kN®u. Пунктир показывает экстраполяцию величин ku®N и kN®u в области излома шеврона. Отклоняющиеся от экстраполяционной прямой точки в верхней левой части графика (т. е. вдали от точки излома, от точки денатурации белка) свидетельствуют либо о какой-то перестройке переходного состояния, либо о появлении каких-то дополнительных метастабильных интермедиатов (возможно, типа расплавленных глобул), которые могут лежать как на, так и вне основного пути сворачивания. Обратите внимание, что все эти перестройки и/или интермедиаты не повышают скорость сворачивания по сравнению с той, что можно было бы ожидать при неизменности переходного состояния: отклоняющиеся точки лежат ниже интерполяционной прямой. Картинка, с небольшими изменениями (добавлены экстраполяционные пунктиры) взята из T. Kiefhaber Proc. Natl. Acad. Sci. USA (1995) 92:.
Действительно, то, что денатурант вообще разворачивает белок, показывает, что он сильнее притягивается к развернутому белку, чем к нативному. То, что денатурант замедляет сворачивание исходно развернутого белка (см. спад скорости в левой части Рис.17-8) показывает, что он сильнее притягивается к исходному, развернутому белку, чем к переходному состоянию; а то, что денатурант ускоряет разворачивание исходно нативного белка (см. подъем скорости в правой части Рис.17-8), — показывает, что он сильнее притягивается к переходному состоянию, чем к исходному, нативному. Это означает, что контакт переходного состояния с денатурантом больше, чем у нативного белка, но меньше, чем у денатурированного. Иначе говоря, приведенный на Рис.17-8 шевронный график показывает, что переходное состояние по степени контакта с растворителем, т. е. по своей компактности находится где-то на полпути между развернутым и нативным состояниями белка. Этот график дает даже больше: так как наклон для kN®u несколько меньше, чем для ku®N, то компактность переходного состояния несколько ближе (в данном случае) к оной у нативного белка, чем у денатурированного.
Рисунок 17-8 относится к лизоциму — белку, денатурация которого гуанидингидрохлоридом приводит прямо к образованию клубка, а не расплавленной глобулы. При умеренных концентрациях денатуранта (при не очень сильном его разбавлении водой) лизоцим сворачивается довольно медленно, за многие минуты. Однако при самоорганизации в почти чистой воде (происходящей при сильном разведении водой крепкого раствора гуанидингидрохлорида, где находились клубкообразные белковые цепи), скорость сворачивания лизоцима максимальна, — и уже слабо зависит от остаточной концентрации денатуранта. При этом в сворачивании лизоцима появляется компактный метастабильный интермедиат типа расплавленной глобулы, — интермедиат, который не наблюдается при повышенных концентрациях денатуранта. Появление компактных метастабильных интермедиатов сворачивания, вообще говоря, всегда "выполаживает" зависимость скорости сворачивания от содержания денатуранта — см. левый край Рис.17-8. Известно, что ренатурация белка, стартующая не от клубка, а от расплавленной глобулы (например, ренатурация карбоксиангидразы), также демонстрирует довольно слабую зависимость скорости ренатурации от концентрации денатуранта. Последнее означает, что переходное состояние в таком сворачивании мало отличается от исходной расплавленной глобулы по компактности.
Иначе говоря, переходное состояние — по "промежуточности" некоторых своих свойств между свойствами нативного и развернутого белка — напоминает расплавленную глобулу. Но это не означает, что переходное — подчеркнем, нестабильное — состояние действительно похоже на расплавленную глобулу. Эксперимент показывает скорее, что переходное состояние можно представить себе (если речь идет о переходе "клубок ® нативный белок") как частично свернутый нативный белок, остальная часть которого все еще находится в денатурированном, клубкообразном состоянии. Вид переходного состояния для переходов типа "расплавленная глобула ® нативный белок" (или "клубок ® расплавленная глобула ® нативный белок") еще не установлен экспериментально, но весьма правдоподобно, что оно включает часть нативной глобулы, в то время как прочая цепь находится в состоянии расплавленной глобулы.
Природу переходного состояния удалось выяснить с помощью белковой инженерии. Применяя множество мутаций, и анализируя соответствующие им изменения в шевронных графиках (см. Рис.17-8, 17-9), удается — ценой колоссального труда — узнать, какие именно остатки вовлечены в "нативоподобную" часть переходного состояния, а какие — нет. Этот метод был разработан А. Ферштом а Англии. Строго говоря, пока что он применяется только к белкам, денатурация которых приводит прямо к образованию клубка, а не расплавленной глобулы.
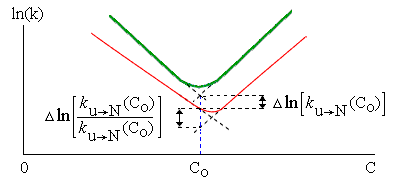
Рис.17-9. Схема, иллюстрирующая сдвиг "шевронного графика" при мутации. Жирная линия: исходный белок; тонкая — мутант. Пунктир показывает экстраполяцию величин ku®N и kN®u в области излома шеврона. С0 — концентрация денатуранта, соответствующая середине денатурационного перехода в исходном белке. На графике показаны измеряемые в этом опыте величины. Одна из них определяет влияние мутации на высоту свободно-энергетического барьера, стоящего на пути из развернутого состояния в нативное: D(F#-Fu) = -RTDln[ku®N]. Вторая — ее влияние на стабильность белка, т. е. на разность свободных энергий нативного и денатурированного состояний: D(FN-Fu) = -RTDln[ku®N/kN®u]. Показанные на графике величины относятся к концентрации денатуранта C0, где требующаяся экстраполяция минимальна (а потому минимальны и погрешности в ней), — но, ценой несколько большей экстраполяции (до C0=0), так обычно определяются изменения в стабильности нативного белка, D(FN-Fu), и в высоте барьера, D(F#-Fu), относящиеся к чистой воде.
Для оценки вовлеченности остатка в переходное состояние (или, как говорят, в "зародыш" сворачивания белка), оценивают, по сдвигу шеврона, влияние мутации данного остатка на (а) скорость сворачивания белка и (б) на его стабильность. Измеряемые величины показаны на Рис.17-9.
Скорость сворачивания белка (перехода U®N) определяется (см. формулу 17.1) разностью свободной энергии "зародыша" (#) и исходного развернутого (U) состояния белка (т. е. величиной F#-Fu).
Если мутация остатка так же влияет на величину F#-Fu, как она влияет на стабильность всего нативного состояния (т. е. на величину FN-Fu), — то это свидетельствует о том, что рассматриваемый остаток так же вовлечен в "зародыш" (образует там те же контакты, имеет ту же конформацию и т. д.), как он вовлечен в нативную глобулу.
Если, наоборот, мутация остатка оказывает влияние только на стабильность белка (т. е. на величину FN-Fu), но не на скорость сворачивания (т. е. она не влияет на величину F#-Fu), — значит, этот остаток не вовлекается в глобулярный "зародыш" сворачивания, т. е. что он входит в нативную белковую глобулу только уже после образования зародыша.
И, наконец, если мутация остатка сильно влияет на стабильность белка и слабее (но с тем же знаком) — на стабильность зародыша, — значит, этот остаток образует в зародыше только часть тех контактов, которые имеет в нативном белке.
Так очерчивается зародыш сворачивания белка (Рис.17-10): для каждого из мутированных остатков цепи вычисляется величина
Ff = D(F#-Fu)/D(FN-Fu) , — | (17.8) |
и, если для данного остатка Ff близка к 1, — то говорят, что он входит в зародыш структуры; а если Ff близка к 0 — то нет.
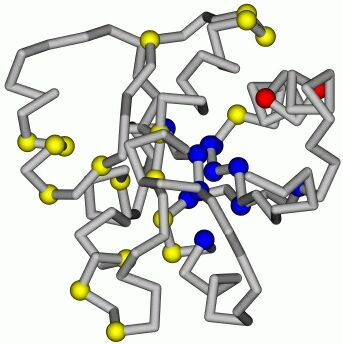
Рис.17-10. Структура переходного состояния белка CheY, согласно T. L![]() pez-Hern
pez-Hern![]() ndes & L. Serrano, Folding & Design (1996) 1:43-55. На фоне нативной укладки цепи CheY синими шариками выделены остатки, вовлеченные в переходное состояние (образующие там более 30% своих контактов), желтыми шариками — не вовлеченные в переходное состояние остатки. Без шариков оставлены те области цепи, где мутации еще не делались. Красным закрашены остатки цепи, сложные для интерпретации, — у них столь малы величины D(F#-Fu) и D(FN-Fu) (последнее важнее, так как эта величина стоит в знаменателе в формуле 17.8), что погрешности в их определении превышают сами эти величины.
ndes & L. Serrano, Folding & Design (1996) 1:43-55. На фоне нативной укладки цепи CheY синими шариками выделены остатки, вовлеченные в переходное состояние (образующие там более 30% своих контактов), желтыми шариками — не вовлеченные в переходное состояние остатки. Без шариков оставлены те области цепи, где мутации еще не делались. Красным закрашены остатки цепи, сложные для интерпретации, — у них столь малы величины D(F#-Fu) и D(FN-Fu) (последнее важнее, так как эта величина стоит в знаменателе в формуле 17.8), что погрешности в их определении превышают сами эти величины.
Замечательно, что лишь очень малое число остатков в белке не удается проинтерпретировать с такой точки зрения (а с нее не удалось бы проинтерпретировать такие остатки, чьи Ff величины лежали бы вне интервала 0 — 1, т. е. те остатки, мутации которых влияли бы только на скорость сворачивания, но не на стабильность нативной структуры; или те, что стабилизировали бы только зародыш, но дестабилизировали бы нативный белок, и т. д.).
Это вселяет уверенность в том, что базовая картина, согласно которой остатки — уж если они вовлечены в зародыш — стоят там так же, как в нативном белке, — эта картина в основном справедлива.
Рисунок 17-10 показывает, что остатки, наиболее существенные для сворачивания белка, группируются в компактный "доменчик", компактное ядрышко сворачивания, причем оно лежит не в геометрическом центре белка, а сдвинуто к его поверхности. Аналогичная картина наблюдается и в других (впрочем, пока немногочисленных) белках, исследованных на предмет местоположения их ядер сворачивания.
В заключение я хочу вернуться к Рис.17-8 и подчеркнуть, что даже для совершенно конкретного белка не существует четко определенного "характерного времени сворачивания". Действительно, этот рисунок показывает, что сворачивание лизоцима занимает порядка 0.1 сек в нативных условиях, и порядка 10000 сек — в условиях созданного гуанидингидрохлоридом (при той же температуре раствора) равновесия нативной и денатурированной форм. А в других условиях, при повышенной температуре, но без гуанидингидрохлорида, области равновесия этих форм отвечает (см. Рис.17-6) время сворачивания порядка 10 сек. Так что, обсуждая скорость сворачивания белка, мы должны иметь в виду либо весь наблюдаемый диапазон времен сворачивания данного белка, либо совершенно конкретные экспериментальные условия — например, те условия, при которых белок сворачивается в клетке.
Лекция 18
Сегодня мы продолжим разговор о самоорганизации белков.
Все экспериментальные данные, о которых я рассказывал, — сколь бы интересны они ни были сами по себе — не дают ответа на вопрос, как белок ухитряется найти свою нативную структуру — среди астрономического числа возможных! — за те немногие секунды или доли секунды, что отпущены на его сворачивание.
А число это — как я говорил, его оценил в еще 1968 г. Сайрус Левинталь — действительно огромно: ~10100 возможных конформаций для цепи из 100 остатков; их "тупой" перебор занял бы ~1080 лет — кладя всего 10-13 секунды на переход из одной конформации в другую.
Как же белок выбирает свою нативную структуру среди бесчисленного множества возможных? — спросил Левинталь, и ответил: — По-видимому, самоорганизующийся белок следует по какому-то специальному "пути сворачивания", и та структура, где этот путь заканчивается, и является его нативной структурой. Иными словами, Левинталь предположил, что нативная структура белка определяется не стабильностью, не термодинамикой, а кинетикой, т. е. она соответствует не глобальному, а просто быстро достижимому минимуму свободной энергии цепи.
Сложность проблемы заключается в том, что поднятый вопрос нельзя решить чисто экспериментально. Действительно: предположим, у белковой цепи есть другая, "ненанивная", но еще более стабильная укладка. Как ее найти, если сам белок ее не находит? Ждать результата в течение 1080 лет?!
С другой стороны, вопрос о том, что — кинетика или термодинамика — определяет укладку белковой цепи, постоянно возникает на пути решения разных прикладных задач. Он возникает, когда речь идет о предсказании структуры белка по его аминокислотной последовательности (надо знать, что предсказывать: самую стабильную или самую быстро сворачивающуюся его структуру). Он возникает и тогда, когда речь идет о дизайне новых, не встречающихся в природе белков (надо знать, что делать: максимально усиливать стабильность желаемой структуры или пролагать максимально быстрый путь к ней).
Однако действительно ли существует противоречие между "структурой стабильной" и "структурой быстро сворачивающейся"? Может быть, стабильная структура автоматически обладает свойством сворачиваться быстро?
Прежде, чем приступить к исследованию этих вопросов, т. е. прежде чем рассматривать кинетические аспекты сворачивания белков, вспомним ряд фундаментальных фактов из области их термодинамики (здесь всюду речь идет об относительно небольших, однодоменных белках, т. е. о белках из 50—200 аминокислотных остатков). Эти факты помогут нам понять, какие условия протекания процесса сворачивания мы должны рассматривать. Термодинамические факты таковы:
1) Разворачивание белка обратимо, причем оно происходит как переход "все-или-ничего". Последнее означает, что в точке денатурации белка только две формы белковой молекулы — "нативная" и "денатурированная" — присутствуют в заметных количествах, а все прочие ("полусвернутые" и "неверно свернутые" формы) практически отсутствуют.
2) Денатурированная форма белков — во всяком случае, небольших белков, развернутых денатурантом, — часто является неупорядоченным клубком.
3) В нормальных физиологических условиях нативная форма белка лишь немногим стабильнее его развернутой формы (а в сам![]() й точке плавления обе эти формы имеют, естественно, одинаковую стабильность). При этом нативная структура белка стабильна благодаря своей низкой энергии, т. е. благодаря сильным взаимодействиям в нативной структуре, а развернутая, — благодаря своей высокой конформационной энтропии, т. е. благодаря огромному числу разных развернутых конформаций. [Необходимое пояснение: как принято в литературе, термин "энергия" здесь означает, строго говоря, всю свободную энергию взаимодействий, в том числе взаимодействий цепи с растворителем (например, "энергия" гидрофобных взаимодействий определяется, как вы должны помнить, энтропией растворителя); термин же "энтропия" здесь охватывает лишь конформационную энтропию цепи, но не энтропию растворителя. Такая терминология принята, чтобы, оставив растворитель за скобками, сосредоточиться на главной проблеме, — как белковая цепь находит "свою" пространственную структуру среди гигантского числа возможных.]
й точке плавления обе эти формы имеют, естественно, одинаковую стабильность). При этом нативная структура белка стабильна благодаря своей низкой энергии, т. е. благодаря сильным взаимодействиям в нативной структуре, а развернутая, — благодаря своей высокой конформационной энтропии, т. е. благодаря огромному числу разных развернутых конформаций. [Необходимое пояснение: как принято в литературе, термин "энергия" здесь означает, строго говоря, всю свободную энергию взаимодействий, в том числе взаимодействий цепи с растворителем (например, "энергия" гидрофобных взаимодействий определяется, как вы должны помнить, энтропией растворителя); термин же "энтропия" здесь охватывает лишь конформационную энтропию цепи, но не энтропию растворителя. Такая терминология принята, чтобы, оставив растворитель за скобками, сосредоточиться на главной проблеме, — как белковая цепь находит "свою" пространственную структуру среди гигантского числа возможных.]
Итак, чтобы разрешить "парадокс Левинталя" и показать, что самую стабильную структуру белковой цепи можно найти за разумное время, мы можем рассматривать только скорость сворачивания этой структуры цепи вблизи точки ее термодинамического перехода, причем перехода типа "все-или-ничего", в клубок ("клубок" — это сумма всех развернутых конформаций белковой цепи). Иными словами, нам достаточно рассмотреть случай, когда самая стабильная укладка цепи лишь чуть-чуть более стабильна, чем клубок, а все прочие формы белковой цепи термодинамически нестабильны. В окрестности этой точки рассматривать сворачивание белка наиболее просто, т. к. здесь нет стабильных интермедиатов сворачивания. Такие интермедиаты, как о том говорилось на прошлой лекции, появляются лишь тогда, когда нативная структура становится много стабильнее клубка. При этом сворачивание белка достигает максимальной скорости, но его анализ усложняется. Поэтому мы ограничимся окрестностью точки равновесия нативной структуры с клубком, где белок сворачивается пусть не максимально быстро, но максимально просто. Причем мы должны рассматривать только такие аминокислотные последовательности, которые обеспечивают наличие большой "щели" между энергией самой стабильной структуры цепи и энергиями всех прочих ее укладок (Рис.18-1): как вы помните, статистическая физика гетерополимеров показывает, что фазовый, типа "все-или-ничего" распад нативной глобулы требует наличия такой щели.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
