
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
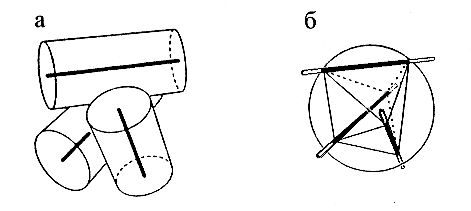
Рис.14-6. Построение многогранника для описания упаковки спиралей. (а) Компактная упаковка трех a-спиралей (цилиндров — 10 ![]() в диаметре — с осями). (б) Построение многогранника: центр упаковки окружается сферой радиусом в 10
в диаметре — с осями). (б) Построение многогранника: центр упаковки окружается сферой радиусом в 10 ![]() ; ее пересечение с осями спирали дает вершины многогранника. На трех из его ребер лежат оси спиралей. Те части ребер, что лежат внутри сферы, оставлены темными. Точки пересечения осей со сферой образуют вершины многогранника. Каждой вершине соответствует одна половинка одной спирали. Оси спиралей образуют часть ребер многогранника, а прочие ребра описывают контакты спиралей.
; ее пересечение с осями спирали дает вершины многогранника. На трех из его ребер лежат оси спиралей. Те части ребер, что лежат внутри сферы, оставлены темными. Точки пересечения осей со сферой образуют вершины многогранника. Каждой вершине соответствует одна половинка одной спирали. Оси спиралей образуют часть ребер многогранника, а прочие ребра описывают контакты спиралей.
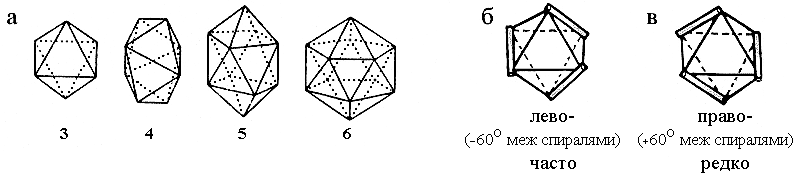
Рис.14-7. Квазисферические многогранники (а), описывающие компактные укладки трех, четырех, пяти и шести спиралей. Большее число спиралей не может уложиться вокруг округлого ядра. Каждый многогранник описывает несколько типов укладок, т. е. типов "штабелей" спиралей, соответствующих различным размещениям осей спиралей на его ребрах. Таких укладок — две для трехспирального комплекса [(б): лево-, и (в): правозакрученный (как на Рис.14-3, 14-4) пучок], десять — для четырехспирального, десять — для пятиспирального, и восемь — для шестиспирального комплекса ("штабеля" для четырех- — шестиспиральных глобул не показаны, но их, при желании, легко построить самостоятельно, разместив спирали — всеми возможными способами — на ребрах многогранника так, чтобы каждая вершина многогранника соответствовала бы одному концу одной спирали). Те упаковки, где межспиральные углы способствуют плотному контакту спиралей — см. Рис.14-9 — встречаются в белках чаще, чем прочие.
Интересно, что в наблюдаемых архитектурах a-спиральных белков вдоль ребер квазисферических многогранников идут не только спирали, но и — как правило — соединяющие их нерегулярные петли (ср. Рис.14-6 и 14-7). Иными словами, в типичном случае белковая цепь как бы обволакивает свое гидрофобное ядро, следуя по непрерывной цепочке ребер квазисферического многогранника.
Обратимся теперь к вопросу о том, как создается плотная упаковка в белковой глобуле. То, что такая упаковка существует, следует из экспериментов, показавших, что белок так же плотен и так же тверд, как органический кристалл. Однако еще предстоит объяснить, как достигается такая упаковка, — слишком уж сложны по форме и разнообразны боковые группы белковой цепи.
Точнее, принцип создания плотной упаковки — и то в самых общих чертах — более или менее ясен только для a-спиралей, почему и уместно рассмотреть этот вопрос именно сейчас.
Первая модель плотной упаковки a-спиралей, упаковка по принципу "выступы (боковые группы) во впадины (между боковыми группами)", была предложена Криком в 1953 г., еще до расшифровки трехмерной структуры первого белка. Затем она была независимо развита Ефимовым и группой Чотиа-Левитт-Ричардсон, и к настоящему времени приобрела вид модели "хребты (боковых групп) в лощины (между таковыми)".
Согласно этой модели, боковые группы на поверхности спирали образуют выступы, создающие разделенные лощинами хребты. "Хребты и лощины" несколько лучше описывают реальность, чем "выступы и впадины", так как разворот одного выступа (одной боковой цепи) в сторону другого (другой боковой цепи) может сделать тот или другой "хребет из выступов" более отчетливым. Хребты (и идущие вдоль них лощины) бывают двух типов. Хребты типа "+4" создаются боковыми группами остатков, расположенных в цепи под номерами "i", "i+4", "i+8", и т. д. (иными словами — с периодом 4), хребты типа "+3" создаются боковыми группами номер "i", "i+3", "i+6", и т. д. (т. е. с периодом 3). Рисунок 14-8 показывает, что эти хребты образуют разного знака углы с осью спирали.

Рис.14-8. a-Спираль; отмечены Сa-атомы (а) и Сb-атомы (б - г). Нумерованные остатки обращены к читателю. Показаны два сорта хребтов (тонкие линии на лицевой поверхности спирали) из сближенных боковых групп (в, г). Хребты из боковых групп "i"-"i+4"-"i+8"... идут под углом -25о к оси спирали (в), хребты из групп "i"-"i+3"-"i+6"... — под углом +45о (г); на рисунке углы представляются меньшими, так как типичные хребты проходят через массивные боковые группы, а на рисунках в, г - через центры Cb-атомов.
При плотной упаковке хребты одной спирали входят в лощины другой. При этом есть две основные возможности:
А) Хребты "+4" одной спирали входят в лощины между такими же хребтами "+4" другой (Рис.14-9а) (здесь для получения плотной упаковки спираль II накладывается на спираль I и поворачивается, пока хребты "+4" обеих спиралей не станут параллельно друг другу). При такой упаковке угол между осями спиралей близок к -500. Этот угол наиболее характерен для контактов спиралей в a-спиральных глобулах. Он также типичен для контакта спиралей в a/b и a+b белках, о которых речь пойдет ниже. Дело в том, что при этом контакте спиралей скрученность слоя a-спиралей (угол скручивания в нем близок к -500/10![]() , где -500 — угол между осями соседних спиралей, а 10
, где -500 — угол между осями соседних спиралей, а 10![]() — ширина a-спирали) неплохо стыкуется с типичной скрученностью b-листа (имеющем тот же угол скручивания, -250/5
— ширина a-спирали) неплохо стыкуется с типичной скрученностью b-листа (имеющем тот же угол скручивания, -250/5![]() , где -250 — угол между осями соседних b-тяжей, а 5
, где -250 — угол между осями соседних b-тяжей, а 5![]() — ширина b-тяжа).
— ширина b-тяжа).
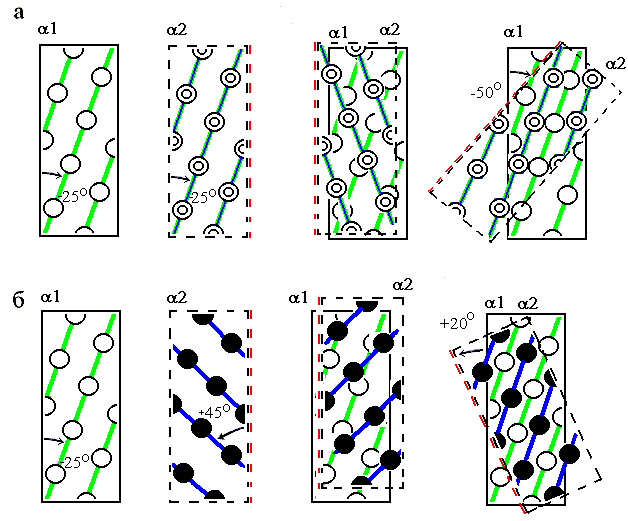
Рис.14-9. Два основных способа плотной упаковки боковых групп при контакте спиралей: под углом -50о (а) и +20о (б). Мы смотрим на зону контакта сквозь одну спираль (сквозь перевернутую вдоль своей оси a2). Остатки "нижней" спирали a1 изображены более светлыми, а верхней (a2) — более темными.
Б) Хребты "+3" одной спирали входят в лощины между хребтами "+4" другой (Рис.14-9б). При такой упаковке угол между осями спиралей близок к +200. Этот угол наиболее характерен для контактов спиралей в пучках — и в a-спиральных глобулах, и в фибриллярных, и в мембранных белках.
Кроме того, хребты "+3" одной спирали могут входить в лощины между такими же хребтами "+3" другой, образуя очень короткий контакт почти перпендикулярных спиралей. В силу малости размера этого контакта, он на рисунке 14-9 не показан, — хотя такой "перпендикулярный" контакт спиралей довольно типичен для a-спиральных глобул.
Заключая описание плотной упаковки, надо заметить, что реальные отклонения от приведенных выше "идеальных" углов весьма велики, так как боковые группы сильно варьируют по размеру. По той же причине в b-структуре (где боковые группы выступают меньше — поверхность b-листа довольно плоская, а спирали — выпуклая) картина типа проникновения хребтов в лощины там совсем смазана и наблюдается лишь в отдельных случаях.
Как согласуется плотная упаковка спиралей с моделью размещения спиралей на ребрах квазисферических многогранников, о которой речь шла выше? Оказывается, довольно любопытным образом. Те "многогранные" упаковки, где углы между спиралями близки к -500 и/или +200, требующимся для плотного контакта спиралей, — эти упаковки встречаются часто; а прочие — редко (но тоже встречаются). Так, из двух изображенных на Рис.14-7 трехспиральных упаковок одна, левозакрученный пучок, приводит к межспиральным углам в -600 (что близко к углу -500, требующемуся для плотной упаковки, см. Рис.14-9а), и такой трехспиральный пучок встречается часто; а другая, правозакрученный пучок, приводит к межспиральным углам в +600 (что далеко от всех углов — -500, +200, 900 — опримальных для плотного контакта), — и такой трехспиральный пучок встречается на порядок реже.
Обратимся теперь к "смешанным" белкам, сложенным из b-листов и a-спиралей. Для них характерна слоистая структура, причем a-спирали и b-участки не могут лежать в одном листе — это привело бы к энергетически невыгодной дегидратации водородных связей на краю b-листа (Рис.14-10).
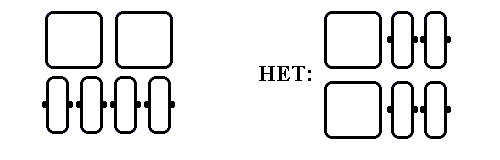
Рис.14-10. Слоистая структура смешанных (a/b, a&b и a+b) белков. Вид с торца b-листа. a-спирали (квадратики - для подчеркивания плотности упаковки глобулы) и b-тяжи (прямоугольники) не могут лежать в одном слое — это привело бы к дегидратации водородных связей (черные точки) на краю b-листа.
Различают a/b ("a дробь b") и a+b ("a плюс b") белки (точнее — домены). Иногда их объединяют в общий разряд a&b ("a и b") белков.
В a/b доменах b-структура параллельна, и a-спирали также параллельны друг другу (и антипараллельны b участкам), а характерное чередование a и b участков в цепи имеет вид - a-b-a-b-a-... .
Есть два характерных мотива строения a/b белков: a/b цилиндр, где b-цилиндр лежит внутри цилиндра, сложенного из a-спиралей (Рис.14-11а), и "укладка Россманна", где более или менее плоский (не считая обычного — правого, если смотреть вдоль b-тяжей — пропеллерного скручивания) b-слой лежит между комплементарно ему скрученными слоями a-спиралей (Рис.14-11б). В отличие от ранее рассмотренных доменов, a/b домены обычно имеют два гидрофобных ядра: в укладке Россманна — между b-листом и каждым из слоев спиралей; в a/b цилиндре — внутри b цилиндра (меньшее ядро) и между b и a цилиндрами (большее).
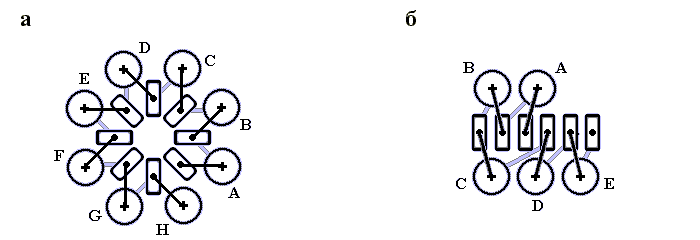
Рис.14-11. Типичные мотивы строения a/b белков и их упрощенные модели (вид на модели — с торца b-слоя): "a/b цилиндр" в триозофосфатизомеразе (а); "укладка Россманна" в NAD-связывающем домене малатдегидрогеназы (б). На детальной картинке в первом белке видна обращенная к нам впадина, образованная расходящимися в форме розетки петлями; она идет к центру b-цилиндра. Во втором — слева вверху видна щель между расходящимися вниз и вверх петлями.
В a/b цилиндре обычно насчитывается восемь a - и восемь b-участков, причем почти все a/b цилиндры имеют одинаковую топологию: все соседние и расположенные через один по цепи b - и a-участки контактируют друг с другом. По-видимому, такое строение обеспечивает особую стабильность белковой глобулы, так как многочисленные белковые глобулы с такой архитектурой (наблюдаемой в 10% белков) все очень похожи друг на друга по форме, — часто и без каких-либо следов общего происхождения в аминокислотной последовательности, и без какой-либо общности функций.
Без общности функций - да; без общности в строении активного центра - да; но не без общности места активного центра в архитектуре глобулы, — каждая архитектура содержит места (обычно — впадины), как бы специально созданные этой архитектурой для активного центра — что бы он ни делал.
Я хочу обратить ваше внимание на "воронку" на оси a/b цилиндра (Рис.14-12а), — вы видите, что эта вмятина, заложенная в общей архитектуре белка, не прикрыта петлями. Здесь находится активный центр. Точнее: из двух таких "воронок", расположенных на противоположных торцах a/b цилиндра, под активный центр используется только одна, — та, куда смотрят С-концы b-участков и N-концы a-спиралей. Считается, что именно эти концы (возможно, из-за множества открытых NH-групп N-концов спиралей), вместе с соединяющими их короткими петлями, особенно пригодны для связывания разнообразных субстратов. Впрочем, этот вопрос еще недостаточно ясен.
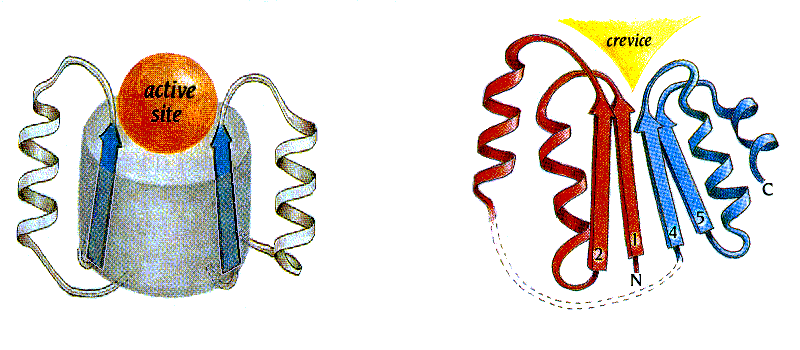
Рис.14-12. Типичное положение активного центра (active site) в a/b белках: в "воронке" на оси a/b цилиндра (а), и в щели (crevice), образованной расходящимися петлями в "укладке Россманна" (б). Картинки, с небольшими изменениями, взяты из [5].
Аналогичное размещение активного центра — в щели, во вмятине, причем во вмятине, куда тоже смотрят С-концы b-участков и N-концы a-спиралей — наблюдается и в "укладках Россманна". Только здесь вмятина образуется не при расхождении петель от центра цилиндра, а при расхождении петель, одна часть которых идет от b-листа к спиралям лежащим под листом, а другая — над этим листом (Рис.14-12б).
Теперь перейдем к a+b белкам. В их основе лежит антипараллельная (а не параллельная, как в a/b белках) b-структура.
В a+b белках выделяются два класса. Белки одного класса (их порой называют ab-складками) напоминают a/b белки тем, что в них слой a-спиралей лежит на b-листе. Они напоминают a/b белки также регулярным (но с иным, чем в a/b белках) чередованием a и b участков в цепи и в пространстве. Белки другого класса ("собственно" a+b белки) не имеют такого чередования, в их цепях a-структура "отмешана" от b-структуры.
Характерное чередование a - и b-участков в цепи ab-складки имеет вид...a-b-b-a-b... или......a-b-b-b-b-a-b-b... (Рис.14-13). Здесь отдельные a-спирали лежат между b-шпильками или b-листами из четного числа b-тяжей. Соседние по цепи b-участки в ab-складках образуют антипараллельные b-шпильки; а из-за четного (а не нечетного, как в a/b белках) числа b-тяжей между a-спиралями, и из-за общей колинеарности этих тяжей a-спиралям, — a-спирали также образуют антипараллельные шпильки. Интересно, что такая (или, точнее, сходная — abbabb) "складчатая" конструкция белка была сначала теоретически предсказана (точнее — предложена в качестве проекта белково-инженерного дизайна), а затем уже такие архитектуры были массово обнаружены в природе, — причем их оказалось особенно много среди РНК-связывающих белков.
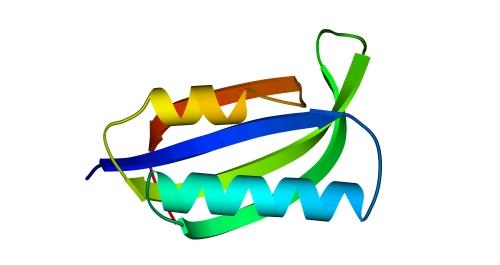
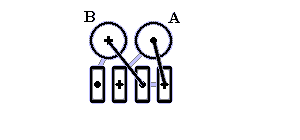
Рис.14-13. Один из типичных мотивов строения a+b белка: "ab складка" (ab-plait) в рибосомальном белке S6. ab складка отличается более регулярным, чем в "собственно" a+b белках, чередованием вторичных структур в цепи (в данном случае: babbab). Радужная кодировка (синий - голубой - зеленый - желтый - оранжевый - красный) позволяет проследить ход цепи, от N - к С-концу. Справа помещена схема строения этого белка (вид с торца структурных участков, приблизительно колинеарных). Спирали занумерованы буквами. "+" на торце a - или b-участка означает, что он идет от нас (т. е. что к нам обращен его N-конец), точка — что к нам.
В "собственно" a+b доменах (Рис.14-14) a - и b-участки расположены в цепи нерегулярно, и скорее — как бы блоками. Такие белки обычно выглядят как b-лист (часто — загнутый сам на себя), прикрытый отдельными a-спиралями или a-спиральным субдоменом; b-структура в них в основном антипараллельна, как в "чистых" b белках.
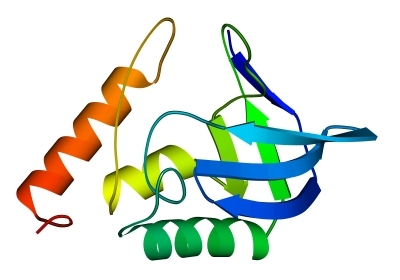
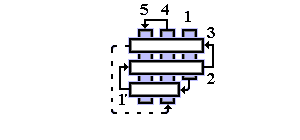
Рис.14-14. Характерный мотив строения a+b белков: нуклеаза стафиллококка. "Собственно" a+b белки отличаются менее регулярным, чем в a/b белках или ab-складках, чередованием вторичных структур в цепи (в данном случае: bbbabbaa). Мотив укладки цепи, наблюдаемый в b-домене нуклеазы, называется "ОБ-укладка" ("OB-fold", то есть "Oligonucleotide-Binding fold", или "укладка [имени] Олега Борисовича [Птицына]"). Справа помещена схема строения этого, часто встречающегося в разных белках, ОБ-домена (вид сверху на ортогональную упаковку b-участков). b-тяжи занумерованы цифрами.
Очень характерной чертой a/b и a+b белков (а равно и b-белков) является правовинтовой (т. е. "к нам — против часовой стрелки") ход перемычек (Рис.14-15) между параллельными b-участками одного и того же b-листа, следующими друг за другом в цепи (но не обязательно при том соседними в этом листе) (см. Рис.14-11 и далее). При этом перемычка между такими параллельными b-участками в a/b - и a+b-белках, как правило, включает в себя a-спираль. В b (и порой в a+b) белках в такой правовинтовой перемычке между параллельными b-участками одного листа лежит — вы должны помнить из прошлой лекции ("abcd" структуры и т. д.) b-участок другого листа, а иногда и отдельный b-лист. Иногда (довольно редко) в перемычке между параллельными b-участками нет ни a-, ни b-структуры, — но и тогда эта перемычка обычно право-, а не левовинтовая.
На следующей лекции мы увидим, что такой ход перемычки позволяет большее разнообразие кодирующих, т. е. стабилизирующих такую структуру последовательностей.
Рис.14-15. Типичный, правовинтовой ход перемычек между параллельными b-тяжами одного листа. В перемычке обычно находится еще один участок вторичной структуры.
Завершая обзор структур глобулярных белков, я хочу еще раз подчеркнуть, что одни и те же — или очень похожие — архитектуры часто встречаются в белках, совсем не сходных функционально или филогенетически. Это открытие заложило основы физической, или рациональной (как это чаще называется в литературе) классификации белков.
Наиболее полная и популярная на сей день классификация представлена в компьютерном классификаторе SCOP (Structural Classification of Protein), сделанном после его переезда из Пущино в Кембридж. Она начинается с класса белка (a, b и т. д.), классы подразделяются по мотивам укладки цепи, и уже мотивы подразделяются на суперсемейства, где просматривается хоть какая-то гомология последовательностей, те — на семейства с явно проявляющейся гомологией, и т. д. Но, пожалуй, еще более последовательно физическая классификация белковых структур проводится классификатором САТН (Class - Architecture - Topology - Homology), сделанном в группе Дж. Торнтон в Лондоне.
Лекция 15
Теперь поговорим об общих закономерностях, наблюдаемых в структуре белков. Из прошлых лекций вы должны были вынести впечатление, что большинство белковых цепей вписывается в узкий набор стандартных структур.
На самом деле здесь действует правило "80%:20%". В его исходном виде оно гласит: "80% всего пива выпивается 20% населения". В применении к белкам — "80% всех белков вписывается в 20% наблюдаемых архитектур белковых глобул". И я позволил себе сосредоточиться именно на типичных структурах.
Вопрос — почему же большинство белков вписывается в узкий набор стандартных структур? И почему не все (как цепи ДНК)? На каком структурном уровне проявляется это сходство? И что стоит за этими общими структурами: память об общем происхождении? функциональная целесообразность? или необходимость удовлетворять общим принципам сворачивания стабильных белковых структур?
По мере роста информации о пространственном строении белковых молекул становилось все яснее, что существуют какие-то "типовые проекты" строения белковых глобул. Архитектуры вновь расшифрованных белков (или, по крайней мере, их доменов) все чаще и чаще оказывались сходными с архитектурами белков уже известных — но при этом совсем других и по функции, и по аминокислотной последовательности. Поэтому причина сходства структур, видимо, заключается не только в эволюционной дивергенции и не (или не только) в функциональной конвергенции белков, а в ограничении набора укладок какими-то физическими закономерностями.
В 70-х годах стало ясно, что между двумя "традиционными" структурными уровнями (вторичная структура белка и его детальная атомная трехмерная структура) находится промежуточный уровень — "мотив укладки" белковой цепи, определяемый взаимным расположением a - и/или b-участков в глобуле — и что именно на этом уровне проявляется сходство белков, не связанных ни эволюционно, ни функционально. В отличие от детальной, атомной трехмерной структуры, "мотивы укладки" удивительно просты и даже красивы (Рис.15-1).
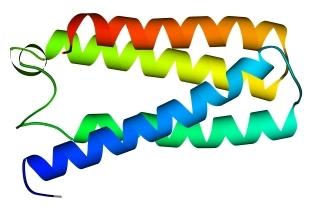


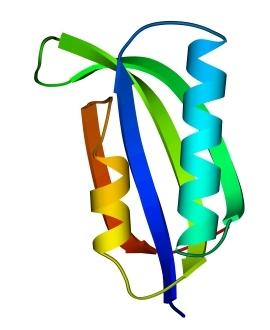
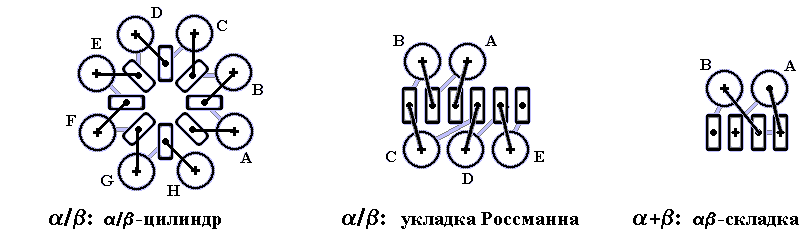
Рис.15-1. Характерные мотивы укладки белковой цепи в a, b, a/b и a+b белках. Внизу — их упрощенные схемы, вид с торца укладки. Обратите внимание на слоевую упаковку a - и b-структур и на то, что каждый слой сложен либо только из a-спиралей, либо только из b-тяжей, но не из a-спиралей и b-тяжей одновременно.
В поисках ответа на вопросы: (1) В чем физическая причина простоты и регулярности типичных мотивов укладки белковой цепи? и (2) Почему одни и те же мотивы встречаются в самых разных белках, и чем замечательны именно эти мотивы? — мы исследуем прежде всего стабильность различных структур. Такой подход оправдывается тем, что одни и те же пространственные структуры белков могут быть получены в результате кинетически совсем разных процессов. Они получаются и in vivo (как в процессе биосинтеза белка на рибосоме, так в процессе транслокации — в более или менее развернутом виде — через мембрану), и in vitro, при сворачивании (ренатурации) целой белковой цепи из развернутого состояния. Это значит, что детальная последовательность действий не играет решающей роли при сворачивании белка.
Начнем с простого вопроса — почему существует слоевое строение глобулярных белков, о котором мы говорили на прошлой лекции. Иными словами, — посмотрим, почему стабильность плотной глобулы требует, чтобы каркас белковой молекулы выглядел бы как компактная упаковка a - и b-слоев, чтобы a - и b-участки шли от одного края глобулы до другого, и чтобы нерегулярные участки не лежали внутри глобулы.
В общем, мы об этом уже говорили. Тут все дело в водородных связях, которые стоят дорого, а потому все должны быть насыщены в стабильной структуре. Доноры и акцепторы таких связей есть в пептидной группе каждого аминокислотного остатка. Насытиться они могут или водой, или при образовании вторичной структуры. Поэтому только вторичные структуры могут не контактировать с водой — лежать внутри глобулы — а содержащие свободные полярные пептидные группы элементы — петли, края b-листов и концы a-спиралей — должны быть на поверхности.
Вытянутые a - и b-структуры должны, ради стабильности глобулы, со всех сторон плотно окружать гидрофобное ядро, создаваемое боковыми группами этих участков, и тем самым отделять его от воды. В то же время a-спирали и b-листы не могут смешиваться в одном слое, — пропадут водородные связи края b-листа. Значит, стабильность глобулы требует образования a-слоев и — отдельно — b-слоев (Рис.15-1). Такие слои (обычно не плоские — скрученные, иногда цилиндрические, а в a-спиральных глобулах — даже квазисферические) действительно, как мы видели, типичны для белковых глобул.
Подавляющее большинство доменов может быть представлено в виде двух-, трех - или (редко) четырехслойных структур, хотя отдельные белки (особенно те, которые содержат металлоорганические комплексы или много S-S связей боковых групп) могут и не вполне удовлетворять этой схеме. Более чем четырехслойных доменов нет — и в принципе ясно, почему. У них внутри, в отдалении от воды находилось бы слишком много остатков, и при типичном для белковых цепей — точнее, для цепей водорастворимых глобулярных белков — соотношении 1:1 между неполярными и полярными остатками многие полярные остатки увлекались бы внутрь белка, что энергетически крайне невыгодно: такой белок не был бы стабильным. Поэтому очень большие единые глобулы "обычного" аминокислотного состава должны быть нестабильны, и большие белки должны разбиваться на субглобулы, домены.
В принципе, можно, видимо, придумать такие аминокислотные последовательности, боковые группы которых как бы "залечат" все разрывы водородных связей между главной цепью и водой, — разрывы, которые последуют за погружением в глобулу края b-листа или петли, так и насытят водородными связями увлеченные внутрь белка полярные боковые группы. Или — придумать последовательности, которые с лихвой заплатят за эти разрывы мощными связями — например, ковалентными (S-S) или координационными (как в металлоорганике). Придумать можно. Но это будут очень специальные, — а значит, очень редкие последовательности...
Может быть, тут-то собака и зарыта, — может быть, "нормальные" глобулярные белки создаются "нормальными" (т. е. сравнительно слабо отобранными), а не "очень редкими" (т. е. сильно отобранными), последовательностями!?
Попробуем взглянуть на первичные структуры белков (Рис.15-2). Статистический анализ показывает, что аминокислотные последовательности водорастворимых глобулярных белков — а о них-то сейчас и идет у нас речь — выглядят как "случайные". То есть в них разные аминокислотные остатки перемешаны примерно так, как можно было бы ожидать при случайной сополимеризации. Конечно, каждая последовательность не есть результат случайного биосинтеза; каждая белковая цепь кодируется геном. Однако аминокислотные последовательности водорастворимых глобулярных белков выглядят как "случайные", — в том смысле, что в них нет ни блочности, характерной для мембранных белков (где явно гидрофобные куски перемежаются с явно гидрофильными), ни периодичности, характерной для белков фибриллярных.
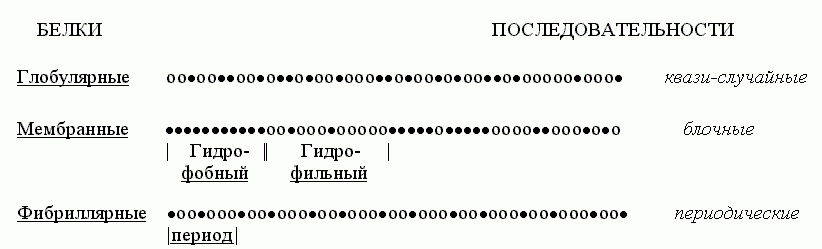
Рис.15-2. Характерные мотивы чередования гидрофобных (·) и полярных (о) аминокислот в первичных структурах водорастворимых глобулярных белков, мембранных белков и фибриллярных белков.
А что такое "выглядеть как случайная последовательность"? Это значит — выглядеть как большинство из всех возможных последовательностей... Значит, рассматривая водорастворимые глобулярные белки, вполне осмысленно ставить вопрос о том, какие стабильные пространственные структуры обычно кодируются самыми массовыми, случайными или похожими на них ("квазислучайными") последовательностями.
Следуя логике такого анализа, один результат мы уже только что получили. Мы выяснили, что типичные упаковки, "штабеля" вторичных структур в глобулярных белках выглядят (Рис.15-1) именно так, как должны выглядеть стабильные упаковки случайных или почти случайных аминокислотных последовательностей.
Пойдем дальше и рассмотрим мотивы укладок белковых цепей.
Как мы уже видели, мотивы укладок белковых цепей часто удивительно красивы. Ход белковых цепей часто напоминает линии, орнаментирующие керамику (Рис.15-3). И, по глубокой мысли Джейн Ричардсон, открывшей это сходство, оно не случайно — так как и линия орнамента, и белковая цепь "решает" одну и ту же задачу — окружить объем (в белке это центр глобулы, ее гидрофобное ядро), избежав самопересечений этой линии.
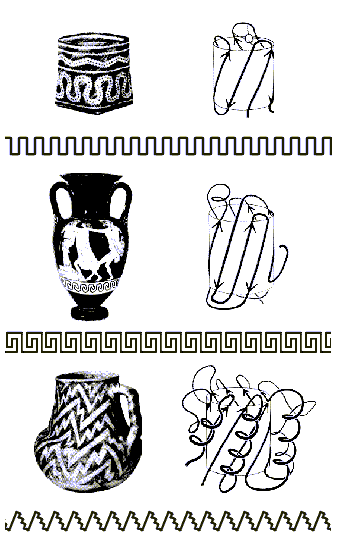
Рис.15-3. Мотивы укладки белковой цепи и орнаменты на индейских и греческих вазах: два решения задачи окружения объема несамопресекающейся линией. Вверху: мотив меандра; в середине: мотив греческого ключа; внизу: мотив зигзага-"молнии". Рисунок взят с обложки Nature, v.268, No.5620, 1977, где была напечатана статья J. Richardson о мотивах укладки белковых цепей.
В белках такой эффект достигается тем, что структурные участки уложены вокруг ядра (или двух ядер; последнее типично для a/b белков), а петли скользят по поверхности ядер — и не перекрывают друг друга (Рис.15-4).
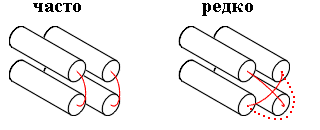
Рис.15-4. Перекрывание петель редко наблюдается в белках, — будь то проход одной петли над другой или обход одной петли вокруг другой.
Чем же плохи перекрывания петель — ведь при этом цепи не врезаются друг в друга, а просто одна из них проходит на другой? Тем, что "нижняя", прижатая к ядру петля лишается водородных связей с водой. А чтобы залечить эту потерю — опять нужны "редкие" последовательности...
Здесь, правда, нас должно смутить то, что при перекрывании петель пропадет лишь одна, максимум две водородные связи — т. е. энергии потеряется немного, килокалорий три или пять. Это не только много меньше, чем полная энергия взаимодействий в белке, измеряемая обычно сотнями килокалорий (судя по опытам по плавлению белков), но и заметно меньше чем обычный "запас стабильности" белка (т. е. разность свободных энергий его нативного и денатурированного состояния). В нативных условиях этот запас составляет — по тем же опытам — порядка 10 ккал/моль. Почему же "дефект" ценой всего в 5 ккал/моль запрещает — или почти запрещает — перекрывание петель в нативных белковых глобулах?
И еще вопрос: что мешает сделать в петле дополнительный изгиб (пунктир на Рис.15-4) — и тем избежать физического перекрывания одной петли другой (т. е. заменить перекрывание петель обходом)? Может быть, здесь дело в упругости полимерной цепи — ведь за дополнительный изгиб петле пришлось бы заплатить (как показывает расчет) несколько (все те же несколько!) ккал/моль?
Здесь любой человек, знающий физику полимеров, должен прервать меня и сказать: "Упругость полимера — энтропийный, а вовсе не энергетический эффект! То есть сильно изогнутая цепь не может флуктуировать так свободно, как прямая или слабо изогнутая. Иными словами, с сильно изогнутой формой цепи совместимо гораздо меньше конформаций, чем с ее более или менее вытянутой формой. Однако эффект, о котором Вы говорите, относится к флуктуирующей цепи, — т. е. к цепи, не фиксированной в глобуле. Но в нативном белке цепь фиксирована, и — так ли она идет, иначе ли — она все равно будет иметь лишь одну какую-то конформацию. Какое отношение имеют описываемые Вами энтропийные потери к нативной структуре белка, где цепь все равно фиксирована, т. е. все равно имеет нулевую энтропию?"
Запомним эти вопросы, а пока рассмотрим еще одну характерную черту белковых архитектур — то, что перемычка между параллельными b-участками почти всегда образует с ними правозакрученную, а не левозакрученную спираль (Рис.15-5).
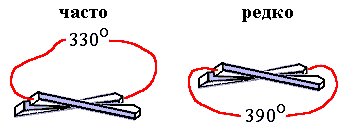
Рис.15-5. Левовинтовой ход перемычек между параллельными b-участками очень редко наблюдается в белках, правовинтовой — часто.
Критерий стабильности позволяет и в этом случае выделить "лучшую" из двух зеркально-симметричных белковых архитектур. В основе различия лежит зеркальная асимметрия природных аминокислот. Она приводит, как вы помните, к преимущественно правому, если смотреть по ходу b-тяжей, скручиванию b-слоев, состоящих из L-аминокислот (это скручивание показано на Рис.15-5). При этим угол между осями соседних b-участков близок к 300, так что полный угол поворота близок к 3300 для правовинтовой перемычки и к 3900 — для левовинтовой. В результате, из-за жесткости полипептидной цепи, правовинтовая — менее закрученная — перемычка выгоднее, чем левовинтовая, т. е. ее свободная энергия ниже, — хотя и опять немного, на пару ккал/моль, и опять в результате не энергетического, а энтропийного эффекта.
И здесь мы опять наталкиваемся на два поставленных выше (и пока оставленных без ответа) вопроса:
(1) Почему "дефект" ценой всего в несколько ккал/моль — на фоне гораздо большей полной энергии белка — может практически запрещать многие мотивы белковых архитектур?
(2) Какое отношение имеют описываемые здесь упругие, т. е. энтропийные эффекты к нативной структуре белка, где цепь все равно фиксирована?

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
