
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
скрещивание инбредных самцов с неинбредными самками другой породы — топкроссбридинг;
скрещивание инбредных самцов одной породы с инбредны-ми самками другой породы — инкроссбридинг;
спаривание инбредных маток с аутбредными самцами — боттомкроссинг
Способы количественного учета и схематического изображения степени инбридинга у потомка разнообразны, но все они в своей основе исходят из данных, характеризующих происхождение, то есть родословную потомка.
Наиболее распространен в практике учет инбридинга, предложенный Шаноружем (1909). По этому методу учитывается число рядов поколений, отделяющих потомка от предка, на которого осуществлен инбридинг. Начиная от ряда родительского поколения, обозначаемого римской цифрой I, далее каждый последующий ряд записывается как П, III, IV и т. д. Повторяющийся предок в родословной потомка со стороны матери и отца записывается с указанием ряда поколений, в которых он присутствует по материнской и отцовской стороне родословной. Например, если потомок (А), или пробанд, произошел от спаривания деда (В) с внучкой (Б) (см. схему), то, по Шапоружу, инбридинг у потомка А на деда В будет записан как III—I, то есть дед В присутствует со стороны матери потомка в III, а со стороны отца в I ряду, то есть имело место близкое родство.
Пробанд А
Мать ББ | Отец В ц В | ||||||
Мать материматери | Отец матери | Мать отца отца | Отец отца отца | ||||
МММ | ОММ | MOM ООМ В | ООМ В | ММО | ОМО | МОО | ООО |
Более точный метод определения степени инбридинга был разработан С. Райт|ом (1921) на основании принципа путевого анализа. Для этого была предложена формула коэффициента инбридинга. Позднее профессором в формулу Райта было внесено уточнение, что делает вычисление более удобным.
Формула Райта — Кисловского представляет коэффициент инбридинга F в виде дроби в границах от 0 до 1 (или в %). В формуле учитывается величина наследственности, получаемой потомком от одного из родителей, равная 0,5, а также число рядов предков по матери — п1 и по отцу — п2, в которых присутствует предок, на которого ведется инбридинг:
![]()
Если потомок инбридирован на нескольких предков, которые, в свою очередь, могут быть инбредными, то формула усложняется.
![]()
где fa — величина коэффициента инбридинга предка, который был сам инбридирован, a Σ — это знак суммирования по всем предкам, на которых заинбридированы потомки
В приведенном выше примере коэффициент инбридинга для пробанда А достигает.
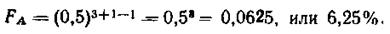
Чем больше величина F приближается к единице (или 100%), тем сильнее инбридирован потомок на предка, тем больше у потомков можно ожидать проявления инбредной депрессии и тем более вероятность повышения гомозиготности потомка по генам предка. Коэффициент инбридинга не указывает в абсолютных цифрах или в процентах, насколько гомозиготен потомок; он только свидетельствует о вероятности того, насколько примененный родственный подбор увеличит гомозиготность потомка по сравнению с исходным состоянием генотипа.
У животных с высокой степенью гетерозиготности инбридинг усиливает гомозиготность значительно быстрее, чем у животных, уже имевших значительную гомозиготность до инбридинга.
Уровень гомозиготности потомка зависит от того, были ли его родители гетерозиготны или гомозиготны.
Так, если отец потомка был гомозиготен и на него проводится инбридинг, то гомозиготность потомков от такого отца резко повышается и уже во втором поколении составляет 0,75 (или 75%), а в третьем — 0,875 (или 87,5%). Если же отец потомка имел гетерозиготный генотип, то при инбридинге, когда он спаривается с дочерью, гомозиготность потомка не повышается, а остается даже при дальнейших спариваниях на прежнем уровне, близком к 0,5, но при этом усиливается генетическое сходство потомков следующих поколений с родоначальником инбредной группы. Использование гетерозиготных производителей — наиболее перспективный вариант инбридинга при проведении племенной работы в стаде, так как он не сопровождается депрессией у потомка, но обеспечивает * более стабильное сходство с ценным предком.
Этот процесс можно проследить по следующей схеме. Допустим, что генотип производителя, на которого планируется вести инбридинг, гетерозиготен по двум локусам (АаВb). При спаривании самца АаВb с самками aabb у потомства первого поколения будут формироваться генотипы, типичные для анализирующего скрещивания в соотношении 1:1:1:1, а именно АаВЬ+Aabb + aaBb + aabb. Из этого следует, что 25% потомков будут иметь гетерозиготный генотип, сходный с предком, то есть АаВb. Если повторить спаривание потомков с тем же гетерозиготным предком АаВb, то у потомков второго поколений гомозиготность останется на том же уровне, что и в первом поколении (то есть 50%), количество потомков, имеющих сходный с предком генотип АаВb, возрастет и составит 39%. Следовательно, доля потомков, сходных по генотипу с отцом, на которого проведен инбридинг, существенно увеличится, что будет соответствовать целям селекции.
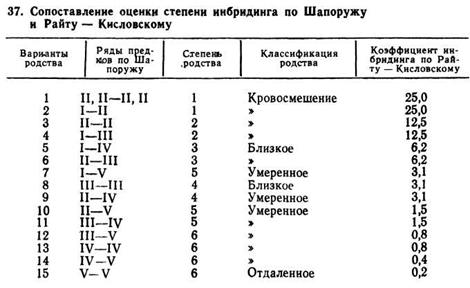
Количественное соотношение степени инбридинга, определенное по Шапоружу и Райту — Кисловскому, приведено в табл. 37.
Наиболее сильное проявление инбредной депрессии можно ожидать с 1-го по 6-й вариант, а с 7-го по 15-й депрессия наблюдается редко или в незначительной степени.
Родство с 7-го по 15-й вариант будет сопровождаться незначительным увеличением гомозиготности, но в данном случае гарантируется генетическое сходство потомка с предком, на которого заложен инбридинг. Это свойство инбридинга имеет практическое значение для племенной работы. В селекционном процессе, направленном на улучшение стада или породы, необходимо осуществить подбор таким образом, чтобы нарастало генетическое сходство потомства с выдающимся предком, что и достигается применением инбридинга.
Если повторяющийся предок встречается только по одной стороне родословной (материнской или отцовской), то инбридинг не ведет к повышению гомозиготности потомка и поэтому формула Райта в данном случае неприменима, Подбор с использованием одностороннего инбридинга усиливает генетическое сходство потомка с предком.
Формула, предложенная Райтом для вычисления степени возрастания генетического сходства между двумя сравниваемыми животными, такова:
![]()
где Rxy — коэффициент генетического сходства между животными х и у; п1 и п2 — число поколений от данных животных до общего предка по женской и мужской стороне родословной; fx и fy — коэффициенты возрастания гомозиготности для животных х и у; fa — тот же коэффициент для общего инбридированного предка.
Величина Rxy указывает на возможное, а не на фактическое возрастание генетического сходства в результате определенного инбридинга.
Коэффициент генетического сходства дочери (или сына) с отцом (или матерью) составляет 50%, так как гаметы каждого из родителей вносят в зиготу половину наследственности. Между внуком (внучкой) и дедом (бабкой) величина R составляет только 25%, между полными сибсами Rxy —50%, а между полусибсами Rxy=25%.
Чем больше поколений включает родственное спаривание, тем выше уровень гомозиготности и инбридинга.
Умеренный инбридинг, повторенный в нескольких поколениях, не сопровождается значительным повышением гомозиготности, но способствует увеличению генетического сходства с выдающимся предком. В качестве примера можно привести потомство известного быка Фаворита шортгорнской породы. Его потомство, полученное через 12 лет после выбытия Фаворита, имело высокое генетическое сходство с ним, и это соответствовало величине RXy = 55,2%, что было следствием длительного инбридинга на Фаворита. Примеров генетического сходства с выдающимся животным достаточно много.
Так, в XVIII в. в Англии братья Коллинги в результате селекций получили быка Комета шортгорнской породы, который был инбридирован на четырех выдающихся предков: быков Фаворита и Фольджамба и коров Феникс и Леди Майнард, при этом корова Леди Майнард и бык Фольджамб были инбредными Животными. Коэффициент инбридинга был достаточно высоким (F = 46,87%), но, несмотря на это, Комет сыграл важную роль в совершенствовании породы. Тесный и многократный инбридинг был применен в России при созданий орловской рысистой породы лошадей, а также в свиноводстве при выведений крупной белой породы.
Депрессия может наблюдаться и при так называемом ложном инбридинге, который является следствием содержания животных в ряде поколений в одинаковых, часто изнеживающих условиях. В целях устранения депрессии при истинном и ложном инбридинге целесообразно периодически содержать животных, намеченных к спариванию, в различных условиях, что способствует формированию некоторого биологического несходства гамет самцов и самок. Этот прием называют интербридингом.
Гетерозис, Родственное спаривание сопровождается инбредной депрессией, повышением гомозиготности инбредного потомства и увеличением генетического сходства потомка с предком. Противоположными биологическими и генетическими свойствами обладает гетерозис.
Под гетерозисом понимают превосходство потомства первого поколения над родительскими формами по жизнеспособности, выносливости, продуктивности, возникающее при скрещивании разных рас, пород животных, зональных типов.
Явление гетерозиса, или «гибридной силы», было замечено в практике животноводства в давние времена, в частности при получении мулов скрещиванием осла с кобылой. Ч. Дарвин впервые дал научное объяснение «гибридной силы», которая возникает у потомства при скрещивании неродственных организмов. Он объяснял этот эффект биологическим несходством мужских и женских гамет, которое вызывается влиянием различий окружающей среды, в которой обитают родители.
Термин «гетерозис» был введен Г. Шеллом (1914), который объяснял наличие «гибридной силы» состоянием гетерозиготности в генотипе организма, формирующейся в результате скрещивания. Гипотеза гетерозиса, сформулированная Г. Шеллом, Е. Истом и X. Хейсом, объясняет явление гетерозиса наличием гетерозиготности различных локусов и проявляющимся при этом сверхдоминированием, то есть когда действие гетерозиготы Аа на проявление фенотипа оказывается сильнее, чем гомозиготного доминантного генотипа АА (то есть эффект действия Аа больше действия АА). Значение гетерозиготности было подтверждено работами , М. Лернера и других ученых.
Другое объяснение гетерозиса, сформулированное Кийблом и Пеллью (1910), основано на том, что при скрещивании организмов, несущих в генотипе разные гомозиготные гены, например ААbb и ааВВ, у помесного потомства рецессивные аллели переходят в гетерозиготную форму генотипа АаВb, при которой устраняется вредное действие рецессивных генов. Влияние доминантных генов на проявление гетерозиса может быть объяснено простым суммарным действием большого количества доминантных генов, то есть имеет место аддитивный эффект.
К. Давенпорт (1908) и Д. Джонс (1917) предложили объяснять гетерозис исходя из гипотезы взаимодействия неаллельных доминантных генов обоих родителей, что дает суммарный эффект, вызывающий гетерозис.
разработал гипотезу облигатной гетерозиготности. Он считал, что в организме имеются полезные (доминантные) и вредные (рецессивные) гены, Если они находятся в гетерозиготном состоянии, то их действие вызывает гетерозис, Если облигатно-гетерозиготные гены находятся в гомозиготном состоянии, то они действуют неблагоприятно Гипотеза облигатной гетерозиготности была, затем развита , В, Я. Борисенко, , Они исходили из эволюционной роли такой гетерозиготности. В процессе эволюции полезные гены сохраняются в гетерозиготном доминантном состоянии» а вредные гены — в рецессивном
Выявлен экологический тип гетерозиса (Меркурьева и сотр., 1980), который вызывается процессом акклиматизации и проявляется у животных первой экологической генерации. Этот тип гетерозиса проявился в повышенной молочности потомства, родившегося в Рязанской области от айрширских коров, завезенных из Финляндии. В последующих поколениях удои снижались до уровня, соответствующего генетическому потенциалу завезенной группы коров.
Современные представления о причинах появления гетерозиса основаны на том, что гетерозис является результатом взаимодействия многих генов. Их множественное действие и приводит к гетерозисному эффекту. Такое объяснение получило название балансового гетерозиса (Добжанский, 1952). В дальнейшем Лернер (1954), (1961—1968) продолжили разработку этого положения. Согласно их утверждениям гетерозис обусловлен действием многих генов, взаимно сбалансированных в геноме в процессе эволюции, которая определяет оптимальное развитие и приспособленность организма к условиям среды.
Если при скрещивании происходит объединение оптимальных геномов обоих родителей, то у потомков первого поколения возникает наиболее благоприятная ситуация в комбинации геномов, что и приводит к проявлению гетерозиса. Следовательно, гетерозиготность, сопутствующая скрещиванию, претерпевает давление различных факторов и тем самым создается сбалансированное взаимодействие генов в геноме,
В практике животноводства иногда наблюдается так называемый отрицательный гетерозис, когда у потомства уровень признака ниже среднего показателя родителей, но несколько выше уровня признака того из родителей, у которого он развит слабее. Чем выше различия в уровне признака родительских форм, тем больше приближается средний уровень признака потомков к уровню признака худшего родителя. Эта особенность в наследовании описана в отношении настрига шерсти у помесей, полученных от скрещивания коз ангорской породы с грубошерстными козами, Настриг шерсти у помесей первого поколения был несколько большим, чем у грубошерстных, но значительно меньше, чем у ангорских коз, у которых он был в 4—5 раз больше по сравнению с грубошерстными и помесными козами.
Для современного животноводства характерно использование скрещивания, сопровождающегося гетерозисным эффектом, особенно для яичного и бройлерного птицеводства. Эта система включает два основных этапа: выведение инбредных линий птицы с применением разных типов инбридинга и скрещивания (кроссирования) линий для получения так называемой гибридной птицы, у которой проявляется гетерозис. Например» в Нидерландах фирма «Еврибрид» работает с двумя кроссами кур яичного направления: «Хайсекс белый» (белая скорлупа, на базе леггорнов) и «Хайсекс коричневый» (при участии род-айланд и нью-гемпшир с коричневой скорлупой). Эти два кросса занимают ведущее положение в мировом яичном птицеводстве.
Работу по созданию гибридной яичной и мясной птицы проводят и в нашей стране. Для осуществления селекции на получение гетерозиса выводят инбредные линии путем спаривания по типу «брат × сестра» в течение 3 — 4 поколений и более, сочетая это с жесткой выбраковкой нежелательных особей. Из большого числа заложенных линий к финалу остается около 10—15% линий при коэффициенте инбридинга в среднем на уровне 37,5% (спаривание полных сибсов в течение трех поколений). Далее скрещивают оставшиеся линии между собой для проверки их на сочетаемость, затем оставляют для производственного кроссирования наиболее удачные сочетания и получают 2-, 3-, 4-линейные гибриды.
Использование эффекта гетерозиса находит применение и в работе с другими видами животных, особенно в мясном скотоводстве, овцеводстве, верблюдоводстве, рыбоводстве. Методы получения эффекта гетерозиса разнообразны. Гетерозис проявляется при межвидовом скрещивании животных: получение мулов от скрещивания осла с кобылой, выведение новых гетерозисных пород путем получения гибридов от скрещивания крупного рогатого скота с зебу (санта-гертруда, бифмастер, чарбрей, бридфорд — в США; сан-пауло — в Бразилии; хауп-голштин — на Ямайке). В нашей стране отдаленная гибридизация проведена между тонкорунными овцами и архаром и выведена новая порода — архаромеринос. В Киргизии и на Алтае получены гибриды яка с симментальским скотом.
Отдаленная гибридизация сопровождается проявлением гетерозиса по ряду хозяйственно ценных признаков.
Проблема получения и усиления эффекта гетерозиса до конца не решена. Основным непреодоленным препятствием является утрата гетерозисного эффекта во втором поколении, то есть гетерозис, полученный в первом поколении, не закрепляется, а утрачивается в последующих поколениях при разведении помесей «в себе». Некоторые методы позволяют поддерживать гетерозис в нескольких поколениях. Одним из наиболее доступных и результативных методов служит переменное скрещивание, применяемое в пользовательном (товарном) животноводстве. При этом из помесей первого поколения, полученных от скрещивания маток породы А с производителями породы В, выделяют лучшую часть маток и скрещивают их с производителем породы С, получают помесей второго поколения, с проявлением гетерозиса при сочетании трех пород (А, В, С). Далее помесей второго поколения можно скрещивать с производителем породы D и получать более сложных помесей, в которых представлена наследственность исходной материнской породы А и наследственность отцовских пород В, С и D. Иных методов, позволяющих сохранить эффект гетерозиса, в животноводстве не разработано.
В практике современного животноводства доказано, что эффект гетерозиса многообразен и выражается в улучшении ценных хозяйственных признаков. Основными показателями гетерозиса являются повышение эмбриональной и постэмбриональной жизнеспособности; снижение затрат корма на единицу продукции; повышение скороспелости, плодовитости, продуктивности; проявление более широких возможностей приспособления к смене условий и новым элементам технологии. Широкий диапазон гетерозисного эффекта, проявляющийся в многообразии реагирующих признаков, является отражением физиологических и биохимических процессов, обусловленных особенностями генетического аппарата гетерозисных животных.
Исследования по выяснению биологических основ гетерозиса проводились в Институте экспериментальной биологии АН Казахской ССР с 1962 г. под руководством академика . Результаты исследований обобщены в монографии (1982), которая может служить дополнительным материалом для понимания гетерозиса и эффекта скрещивания. В процессе работы определено количество ДНК, РНК, белков и активность ряда ферментов в тканях и в субклеточных структурах клеток (ядра, хромосом) чистопородных и помесных овец. Были выявлены особенности обменных процессов и гетерозис у животных, различающихся по происхождению. Оказалось, что гетерозисный эффект не связан с изменением количества наследственного вещества в отдельно взятой клетке, ядре или хромосомах. Скрещивание не вызывает у помесей активацию ранее неактивных генов, полученных через хромосомы родителей, и не приводит к коренной перестройке обменных процессов. Вместо этого наблюдается лишь стимуляция уровня напряженности метаболических процессов. В процессе онтогенеза это напряжение снижается и уменьшается эффект гетерозиса у помесей.
Биохимический эффект гетерозиса у помесей проявился в стимуляции активности тканевых ферментов (ДНК-азы, РНК-азы и др.), которые влияют на синтез нуклеиновых кислот. Активность ферментов у помесей протекает в более широком диапазоне рН среды, что повышает экологическую пластичность помесных организмов и приспособленность к условиям среды. Следовательно, скрещивание влияет на механизм регуляции активности ферментов.
Синтез РНК в клеточном ядре и трансляция направляемого РНК синтеза молекул белка в цитоплазме протекают у помесей на более высоком уровне. Этому способствует обогащение ядер клеток негистоновыми белками хроматина, который является специфическим стимулятором активности генома. Следовательно, скрещивание стимулировало синтез рибосомальной РНК, то есть усилило процесс транскрипции. Высказывается гипотеза, что с помощью биологически активных веществ (гормонов, метаболитов), которые могут влиять на активность генетического аппарата, можно продлить действие гетерозиса в течение более продолжительного отрезка онтогенеза.
Существуют и другие биохимические объяснения гетерозиса. Считается, что главной причиной гибридной мощности служит формирование на хромосомах чувствительных копий структурных генов, которые образуют избыток информации в клетках и определяют высокую сочетаемость процессов метаболизма (Северин, 1967).
Объяснения гетерозисному эффекту можно найти в суждениях, что у помесей присутствуют полиморфные типы белков (изоферменты), которые различаются некоторыми свойствами.
У родительских форм отсутствует полиморфизм ферментов, а при их скрещивании у помесей формируется полиморфизм и число полиморфных локусов у них поэтому больше, чем у родителей. Это, по мнению некоторых ученых (Финчем, 1968; Кирпичников, 1974), объясняет эффект сверхдоминирования. (1975) считает, что взаимное стимулирование геномов при оплодотворении равносильно аддитивному эффекту объединенных генетических систем и является основой появления гетерозиса, но не является причиной возникновения новых качеств в генетическом материале, поэтому гетерозис проявляется в количествнных изменениях признаков и имеет полигенный тип наследования.
Новый подход в объяснении гетерозисного эффекта предлагает (1968). Он считает, что гетерозис имеет биофизическую основу, так как при оплодотворении происходит обмен электрическими зарядами гомологичных хромосом, что повышает активность хромосом в гибридных зиготах. Это приводит к накоплению кислых белков и РНК, повышает ядрышко-ядерное соотношение и увеличивает скорость митотического деления.
Приведенные объяснения причин гетерозисного эффекта указывают на отсутствие единства в научном объяснении явления гетерозиса, и поэтому проблема остается для дальнейшего изучения и рассмотрения. Несмотря на это, в практике животноводства осуществляют приемы селекции животных на закрепление и усиление эффекта гетерозиса. Существует несколько приемов для вычисления величины эффекта гетерозиса. Выделяют так называемый истинный тип гетерозиса, который определяется по величине превосходства признака у помесных животных над обоими родительскими формами. Другой тип гетерозиса — гипотетический, когда признаки помесного потомства превосходят среднеарифметический уровень признака обоих родителей.
Если отсутствуют данные по одной из пород, от которых получены помеси, то их показатели сравнивают с материнской породой, а улучшенные показатели помесей называют не гетерозисом, а эффектом скрещивания.
Обобщая современное понимание явлений инбредной депрессии и гетерозиса, можно сделать выводы о необходимости использования обоих явлений в практической племенной работе.
Контрольные вопросы. 1. В чем проявляются биологические и генетические свойства инбридинга? 2. Методы оценки степени инбридинга. 3. Суть теорий, объясняющих явление инбредной депрессии. 4. Типы инбридинга, используемые в животноводстве; их терминологические названия. 5. Теории, объясняющие гетерозис. 6. Как используют явление гетерозиса в практике животноводства?
ГЛАВА 15. ГЕНЕТИКА ИММУНИТЕТА, АНОМАЛИИ И БОЛЕЗНЕЙ
В составе современной биологии иммунология оформилась и развивается как самостоятельная наука. Объектом ее изучения служит способность организмов проявлять иммунитет, то есть реагировать в виде неспецифической или специфической реакции на воздействие чужеродных для него веществ (антигенов), проникших в организм. В основе иммунологических методов исследований положено определение реакции клеточных элементов иммунной системы — лейкоцитов, которые синтезируют защитные белковые вещества — антитела — или осуществляют разрушение чужеродных элементов. В основе процесса иммунной защиты лежат генетические особенности клеток защитной системы. В связи с этим в иммунологии интенсивно осуществляется разработка генетических основ иммунитета, выявляется генетика элементов защиты и биосинтеза антител (то есть иммуноглобулинов).
Краткая история развития иммунологии. Потребность защищать людей и животных от болезней сопровождалась поисками эффективных приемов. В Англии в 1774 г. для предупреждения заболевания человека оспой применили обработку кожи человека веществами из пустул коровы, болеющей оспой.
Научно обоснованные исследования были проведены (1893), открывшим явление клеточного иммунитета в виде фагоцитоза, и Луи Пастером, применившим вакцинацию против бешенства, холеры кур, рожи свиней, сибирской язвы. Эти ученые заложили основу современной науки по иммунологии.
В 1897 г. П. Эрлих (1897) разработал принципы биохимического подхода в понимании процесса иммунитета, основываясь на особенностях строения антител. Был открыт такой элемент защиты, как комплемент (Борде, 1899). Наука об иммунитете бурно развивалась в первые десятилетия двадцатого века.
В дальнейшем с применением методов электрофореза, иммунодиффузии, позволивших выявлять качественные различия в белковых компонентах, появилась возможность определять и количественные их уровни. В период 1938—1953 гг. с помощью этих методов выявлена структура антител и иммуноглобулинов (Потер, 1958), определена аминокислотная последовательность иммуноглобулинов разных типов (Эдельман, 1959).
Были выявлены патологические иммунные процессы, такие как явление аллергии (Пирка, 1905), иммунологическая толерантность (Медавар, 1958), явление тканевой антигенной гистосовместимости (Доссе, 1958), аутоиммунные антитела и др. Появились доказательства участия некоторых генов в синтезе иммунных веществ.
Современная иммунология направлена на выявление механизмов иммунного ответа и его генетической обусловленности. Осуществляются поиски целенаправленного воздействия на иммунный ответ организма и возможность его регуляции в целях борьбы или профилактики заболевания.
Иммунология приобрела не только практическое значение для медицины и ветеринарии; она в значительной степени становится необходимым элементом, создающим научное обоснование для животноводства, в частности в биотехнологии воспроизведения и при проверке правильности записей о происхождении животных. Так, например, выяснена иммунологическая основа процесса оплодотворения и обеспечения нормального эмбрионального развития потомства. Установлены иммунологические причины патологии этих процессов. Показано, что для нормального постэмбрионального развития животного важное значение имеет генетическая обусловленность иммунологического статуса организма, которая определяется наследственностью его родителей. В формировании иммунного статуса важную роль играют многие биохимические вещества (например, гормоны и ферменты), которые несут иммунную информацию и влияют на ход развития организма. Эти вещества также имеют генетическую обусловленность.
В условиях современной интенсивной технологии воспроизведения потомства успех такого биотехнологического приема, как пересадка эмбрионов от высокоценных родителей-доноров в организм самки-реципиента, в значительной мере определяется иммунными взаимоотношениями всех участников в создании потомка, а именно: иммунными взаимоотношениями гамет отца и матери при оплодотворении, воздействием матери на зарождающийся эмбрион, взаимодействием трансплантированного эмбриона с организмом самки-реципиента. Поэтому знание иммунных процессов, определение иммунного статуса всех участников биотехнического комплекса в интенсификации воспроизведения будут определять эффект применяемого на практике метода пересадки эмбрионов.
Защитная функция. Различают две основные формы защитной функции: неспецифическую и специфическую, каждая из которых имеет определенные особенности в защите организма. Вместе с тем между обеими формами защиты организма существует взаимодействие.
Неспецифическую функцию защиты осуществляют кожа, слизистые оболочки и их выделения. Дополнительная и важная роль принадлежит реакции фагоцитоза, при которой специализированные клетки (нейтрофилы, моноциты, макрофаги) уничтожают внедряющихся микробов, Гуморальными факторами неспецифической защиты служат белковые (ферментные) вещества: естественные иммуноглобулины, лизоцим, интерферон, бета-лизий, пропердин, комплемент и др. Эта группа веществ определенным образом воздействует на источник инфекций.
Специфическую функцию осуществляют так называемые естественные (нормальные) антитела или иммуноглобулины. При уменьщении их количества снижается резнстентность к инфекциям. Естественные антитела являются ответом иммунной системы на проникновение неизвестного антигена, но проявляют специфическую реакцию. Специфическая защита проявляется как реакция организма в виде синтеза специфических веществ (антител) на проникшие в организм антигены, имеющие различную природу. Специфика этой защиты проявляется 3 том, что на каждый конкретный антиген лимфоциты типа В синтезируют в организме специфическое антитело и между ними происходит взаимная реакция, при которой образуется комплекс антиген — антитело, приводящий к устранению антигена. В рассматриваемом случае реакция может быть выражена различным образом: в виде преципитации, агглютинации или гемолиза.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
