
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У животных сигнальными веществами являются различные гормоны, В клетках вырабатывается специфический белок-рецептор, обладающий способностью связываться с гормоном, который приобретает свойство индуцировать работу одного или нескольких генов, Наиболее изучен этот процесс для половых гормонов, В настоящее время обнаружены и другие сигнальные вещества, так называемые эмбриональные индукторы, а также вещества, попадающие в организм извне.
У эукариот возможна регуляция синтеза белков на уровне трансляции, При этом имеют значение типы тРНК и ферментов, активирующих соответствующие аминокислоты, а также вырожденность генетического кода. Большая часть аминокислот кодируется несколькими кодонами, получившими названия изоакцепторных кодонов. Одна и та же аминокислота может доставляться на мРНК несколькими типами тРНК. Так, кодирование аминокислоты лейцина может происходить посредством кодонов ЦУЦ, ЦУУ, ЦУГ. Процесс трансляции зависит также от состояния тРНК, рибосом, наличия или отсутствия соответствующих ферментов, в том числе и способных модифицировать готовые белковые молекулы.
Из приведенного следует, что молекула ДНК в процессе биосинтеза осуществляет реализацию наследственной информации. Этот процесс, несмотря на некоторые особенности, характерные для прокариот и эукариот, является, по сути, единым для всего органического мира путем воплощения наследственной информации в свойства и признаки.
Современное представление о гене как единице наследственности. В представлении Г. Менделя единицей наследственности был фактор, контролирующий проявление в доминантном или рецессивном состоянии одного признака. В дальнейшем понятия о гене были развиты в работах Т. Моргана, который показал, что ген — это локус (участок) хромосомы, занимающий в ней строго определенное положение.
В современном понимании ген — это функциональная единица молекулы ДНК, контролирующая последовательность аминокислот в кодируемой полипептидной цепи. Специфичность гена определяется числом нуклеотидов и их уникальной последовательностью. Ген имеет определенную величину, выраженную числом нуклеотидов и молекулярной массой. Ген, кодирующий синтез полипептидной цепи, называется структурным. Он является составной частью оперона, имеет сложную систему регуляции, осуществляемой акцепторными генами. Для каждого структурного гена характерна уникальная последовательность нуклеотидов, позволяющая его идентифицировать.
Структурный ген является дискретной целостной единицей, кодирующей синтез одной полипептидной цепи. Любое изменение порядка чередования нуклеотидов — выпадение, добавление или замена хотя бы одного нуклеотида — инактивирует структурный ген или изменяет его функцию.
Ранее отмечено, что для структурных генов эукариот характерно мозаичное строение (рис. 40): участки молекулы ДНК, кодирующие аминокислоты в полипептидной цепи, — экзоны (кДНК) чередуются с участками, которые не обладают этой способностью, — нитронами.
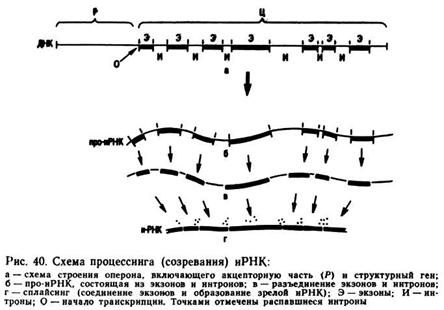
Акцепторные гены каждого оперона обладают высокой специфичностью — к ним могут присоединяться только определенные молекулы белка, в том числе белок-репрессор, подавляющий активность структурных генов, Cap-белок, а также ферментативное белки, обеспечивающие репликацию и транскрипцию. Доля структурных и акцепторных генов в общей ДНК в геномах разных организмов колеблется от 98 до 15%. Остальная часть ДНК генома получила название избыточной ДНК. Особенно много избыточной ДНК содержится 8 геномах растений. Для избыточной ДНК характерно наличие повторов — одинаковых последовательностей нуклеотидов. Р. Бриттен и Д. Кон в 1968 г. установили, что у мыши 70% ДНК составляют уникальные последовательности нуклеотидов, а 30% — повторы; у человека — 66% уникальные последовательности, а 34% — повторы.
Повторы ДНК у эукариот могут иметь различную природу. Некоторые структурные гены, имеющие уникальную последовательность нуклеотидов, могут быть представлены несколькими копиями. Гены, кодирующие гистоны — основные белки, входящие в состав хромосом, в молекуле ДНК представлены различным числом копий, например, в гаплоидном геноме мыши содержится 30 структурных генов, кодирующих гистон Н4. У животных имеются повторы структурных генов, кодирующих глобин, иммуноглобулин, интерферон и другие жизненно важные молекулы белка. Среди повторов генов имеются нефункционирующие гены, которые из-за выпадения или добавления нуклеоти-да потеряли способность синтезировать мРНК. Их называют псевдогенами.
Особенно многократно в молекуле ДНК встречаются повторы структурных генов, контролирующих синтез рибосомальной и транспортной РНК. Так, в гаплоидном геноме лягушки имеется около 8000 генов тРНК, в геноме курицы — около 100 генов рРНК, в геноме дрозофилы их около 130.
В ДНК геномов содержатся и другого рода повторы. Они представляют собой короткие последовательности нуклеотидов, каждый из них содержит около 300 нуклеотидных пар, а также 40000—80000 повторов В1, содержащих приблизительно по 140 нуклеотидных пар.
В составе избыточной ДНК у эукариот в довольно большом количестве содержатся последовательности нуклеотидов, генетическая роль которых пока еще остается невыясненной. Они получили название сателлитной ДНК, которая представляет собой последовательности, состоящие из нескольким нуклеотидных пар. У мыши они состоят из 6 пар нуклеотидов, в том числе 5 пар AT и пары ЦГ; у морской свинки сателлитная ДНК состоит из 6 пар нуклеотидов, в том числе 3 пар ЦГ и 3 пар ТА, АГ и AT. Блоки (кластеры) сателлитной ДНК преимущественно сосредоточены в гетерохроматиновых районах хррмосом, расположенных около центромеры.
Транспозоны. В течение длительного времени считалось, что положение генов в хромосоме и, следовательно, в молекуле ДНК является строго фиксированным, хотя Б. Мак-Клинток еще в 1953 г. доказала, что в геноме кукурузы содержатся так называемые подвижные генетические элементы. В 1975—1977 гг. советский ученый обнаружил в геноме дрозофилы гены, представленные десятками копий и рассеянные по разным хромосомам. Им было установлено, что эти гены являются подвижными или «прыгающими», так как могут быть локализованы у разных линий и даже у отдельных особей в разных хромосомах и в разных локусах одной хромосомы.
Перемещение фрагмента ДНК, содержащего ген или гены, из одной хромосомы в другую, им несвойственную, называется транспозицией. Фрагменты ДНК, способные перемещаться из одной хромосомы в другую или из одного локуса в другой называют транспозонами. Транспозиция включает два процесса: эксцизию и инсерцию. Эксцизией называется освобождение транспозона из молекулы ДНК, в которую он был встроен, а инсерцией — процесс встраивания транспозона в новый локус ДНК.
Транспозоны условно можно разделить на несколько классов. Один из них, наиболее изученный и широко представленный, обнаружил у дрозофилы и назвал их «мобильные диспергированные гены» (МДГ). У дрозофилы имеется около 20 семейств таких МДГ, каждое из которых содержит от 10 до 150 копий, локализация которых в геноме сильно варьирует. Характерной особенностью МДГ являются одинаково ориентированные длинные концевые повторы (ДКП). ДНК МДГ содержит 5—10 тыс. нуклеотидных пар, в том числе 250—1500 нуклеотидных пар — это ДКП. Образование большого числа копий МДГ происходит следующим образом: на матрице ДНК в локусе МДГ-элемента синтезируется РНК, на которой при участии фермента обратной транскриптазы образуется много копий фрагментов ДНК, соответствующих МДГ, которые внедряются в новые локусы ДНК генома. В ДКП МДГ-элементов имеются сигнальные последовательности для начала и окончания транскрипции, а также усилители (энхансеры), резко увеличивающие интенсивность транскрипции. Они содержат также оперон, кодирующий обратную транскриптазу.
Другой класс активных транспозонов (МДГ) включает последовательности ДНК, кодирующие фермент транспозазу, который отвечает за транспозицию МДГ — вырезание и встраивание транспозонов. К ним относят хорошо изученные Р-элемент дрозофилы и Ас-элемент кукурузы.
К особому классу можно отнести пассивные транспозоны — фрагменты ДНК, которые ничего не кодируют, но многочисленные копии которых могут служить субстратом для транспозазы. К их числу могут быть отнесены и длинные обращенные повторы, и даже некоторые МДГ-элементы.
К транспозонам относят также и другие участки генома, если они активно синтезируют РНК, а затем при участии фермента ревертазы образуют многочисленные копии ДНК, кото-рые вставляются в различные участки генома. К их числу относятся два класса коротких транспозонов мыши, названные В1 и В2; В1 содержит 130, В2— 190 пар нуклеотидов. Они рассеяны по всему геному, и почти в каждом фрагменте ДНК содержится В1 или В2, или оба. Находясь в ДНК генома, они активно транскрибируют РНК, а затем при участии обратной транскриптазы образуют огромное число копий (в клетке может содержаться до копий каждого транспозона).
В геноме человека также обнаружены В1 и В2, а также транспозон А1и содержащий 300 пар нуклеотидов и представленный копиями.
Транспозиция играет значительную роль в реализации наследственной информации и может быть причиной наследственного изменения признака (мутации). Многие транспозоны служат матрицами для транскрипции мРНК, кодирующей различные ферменты, в том числе обратную транскриптазу. Внедряясь в новые локусы генетического аппарата клетки, транспозоны влияют на работу окружающих генов. Иногда внедрившийся транспозон изменяет структуру гена вплоть до создания нового, несвойственного данному локусу. Транспозоны могут вызвать глубокие перестройки генома, в том числе делеции, инверсии, транслокации. Для разных генетических локусов от 10 до 90% всех спонтанных мутаций являются результатом транспозиции МГД.
В обычных условиях транспозиция происходит весьма редко, но под действием некоторых факторов наблюдаются так называемые транспозиционные взрывы, когда в клетке сразу перемещается большое число транспозонов, относящихся к разным классам.
В последние годы установлено, что транспозиция и образование большого числа повторов МДГ сходны с ретровирусами птиц и млекопитающих. Ретровирусами называют вирусы; у которых генетическая информация записана на РНК (РНК-содержащие вирусы). Когда такой РНК-содержащий вирус проникает в клетку, при участии фермента обратной транскриптазы синтезируются ДНК-копии РНК вируса. ДНК внедряется в различные локусы генома клетки и становится составной частью молекулы ДНК. Такую ДНК называют провирусом. В геноме мыши может содержаться несколько семейств провирусов, локализованных в разных локусах ДНК. На этих ДНК может синтезироваться РНК и даже могут образовываться вирусоподобные частицы, но инфекционный вирус не возникает. Вирусы, информация о которых содержится в ДНК высших организмов, получили название эндогенных вирусов (ЭВ), а кодирующие их генетические элементы — эндогенных провирусов (ЭП). Так, у кур выявлено 29 локусов ЭП, которые встречаются в различных сочетаниях и с разной частотой. Ни один из локусов ЭП не является обязательным элементом генома кур.
Подавляющее большинство ЭП дефектны и не могут кодировать вирионы, поэтому они не являются инфекционными для родительских клеток. Вместе с тем некоторые ЭП следует рассматривать как генетические факторы риска, повышающие вероятность начала канцерогенного процесса или появления нового онкогенного вируса.
Контрольные вопросы. 1. Каким образом ДНК сохраняет, передает и реализует наследственную информацию? 2. Какую функцию выполняет каждый входящий в оперон фрагмент ДНК? 3. Свойства генетического кода 4. Что такое «ген» в современном понятии? Функции и свойства генов различных типов. 5. Как осуществляется синтез и выделение генов? 6. Народнохозяйственное значение генетической инженерии.
ГЛАВА 8. БИОТЕХНОЛОГИЯ
И ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ
Термин «биотехнология» появился в середине 70-х годов в связи с успехами в области генетической инженерии, биохимии, микробиологии и других смежных областей биологической науки. Современная биотехнология представляет собой новую форму промышленной технологии, основу которой составляют биологические объекты — животные, растения, ткани различных органов, соматические клетки, размножаемые вне организма, микроорганизмы — бактерии, грибы. В основе биотехнологии — генетическая инженерия.
Генетической инженерией называют область молекулярной генетики, разрабатывающую методы конструирования новых функционально активных генетических программ. Датой зарождения генетической инженерии принято считать 1972 г., когда П. Берг с сотрудниками (США) создали первую реком-бинантную молекулу ДНК. Она состояла из фрагмента ДНК, взятого у обезьяньего вируса ОВ40, и бактериофага λ с галактозным опероном Е. colt. Важную роль в генетической инженерии играют ферменты, с помощью которых можно получать определенные фрагменты ДНК исшивать их, например рестриктазы (рестригирующие эндонуклеазы) и лигазы, которые лишены видовой специфичности, поэтому можно получать фрагменты ДНК и сшивать их независимо от того, из одного или разных организмов они выделены. Большое значение для развития генетической инженерии имеет метод секвенирования (расшифровки) первичной структуры ДНК, разработанный в 1977 г. Ф. Сенджером и У. Гильбертом. Этот метод позволяет определить последовательность нуклеотидов в молекуле ДНК с предельным разрешением в один нуклеотид.
Генетическая инженерия как метод конструирования генетических программ включает ряд сложных приемов, объединяющих усилия биохимиков, генетиков, микробиологов:
синтез или выделение соответствующего гена (или генов);
включение данного гена в вектор, обеспечивающий его размножение (клонирование);
трансгеноз — перенос гена с помощью вектора в клетку-рециент и включение в ее геном;
функционирование гена в клетке-реципиенте (адаптация гена).
Синтез генов. Впервые химический синтез гена осуществил в 1969 г. работающий в США индийский ученый X. Г. Корана с сотрудниками. Он синтезировал ген (участок молекулы ДНК), кодирующий синтез аланиновой тРНК пекарских дрожжей. Этот ген состоит из 77 пар нуклеотидов, последовательность которых была известна. Вначале синтезировали мелкие фрагменты ДНК, содержащие от 4 до 13 нуклеотидных пар, затем с помощью фермента лигазы их соединили в соответствующем порядке, но данный ген был не в состоянии синтезировать аланиновую тРНК так как не содержал акцепторной системы.
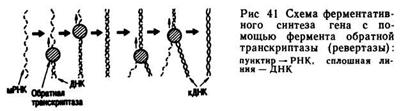
В 1976 г. в лаборатории X. Г. Кораны был синтезирован фрагмент ДНК, включающий структурный ген супрессорной тирозиновой тРНК длиной 126 пар нуклеотидов, промотор, состоящий из 52 пар нуклеотидов, и терминатор, имеющий 21 пару нуклеотидов. К концам молекулы ДНК были прикреплены так называемые «липкие концы», состоящие из чередования нуклеотидов ААТТ (один конец) и ТТАА (другой конец). Благодаря этому данный ген был встроен в геном фага и нормально в нем функционировал. Таким образом, была показана возможность искусственного синтеза генов. Метод позволяет синтезировать относительно мелкие гены, содержащие небольшое число пар нуклеотидов. Гены, кодирующие ферменты или структурные белки, состоящие из тысячи и более нуклеотидных пар, рациональнее создавать методом ферментативного синтеза с помощью фермента обратной транскриптазы (ревертазы). С помощью данного фермента с мРНК могут быть получены точные копии ДНК (кДНК).
Ферментативный синтез может быть схематично представлен следующим образом. В пробирку, содержащую физиологическую бесклеточную среду, вносят дезоксинуклеотидтрифосфаты всех четырех типов (А, Г, Т, Ц), фермент ревертазу, мРИК, кодированную природным геном, копию которого планируется получить. В качестве «затравки», ускоряющей реакцию, вносят небольшие участки молекулы ДНК, содержащие 8 — 10 повторов тимина. На мРНК обратная транскриптаза синтезирует комплементарную ей нить ДНК. Затем на синтезированной нити ДНК строится вторая комплементарная нить ДНК. В результате получают фрагмент двойной спирали ДНК — точную копию того гена, с которого была транскрибирована мРНК (рис. 41).
Описанным способом были синтезированы гены, кодирующие глобины человека, кролика, мыши, утки, голубя, иммуноглобулин мыши, белок хрусталика глаза быка, яичный белок и другие. Подобным образом можно синтезировать структурные гены, не содержащие регуляторную часть оперона, что ограничивает их функционирование.
Выделение генов. Впервые выделить ген методом трансдукции удалось Дж. Бексвиту с сотрудниками. Они показали, что бактериофаг λ при размножении в клетке Е. coli может захватить и встроить в свой геном полный лактозный оперон бактерии вместе с примыкающим к нему геном-регулятором. Фрагмент ДНК Е. coli встраивается 8 геном фага к в строгом порядке: структурные гены z, а, у, оператор — о, промотор р и регулятор i. Применением денатурации и центрифугирования из ДНК-фага был выделен лактозный оперон и ген-регулятор E. coli. К сожалению, метод оказался сугубо специфичным для данного гена и не может широко использоваться в генетической инженерии. В настоящее время генетическая инженерия использует методы, позволяющие одновременно выделить из молекулы ДНК фрагмент соответствующего гена и встроить его в вектор, посредством которого он может быть размножен и включен в геном клетки-реципиента.
Фрагменты ДНК, содержащие ген, чаще всего получают с помощью ферментов — рестриктаз. Эти ферменты разрезают молекулу ДНК в строго определенном месте, где находятся нуклеотиды, распознаваемые данной рестриктазой. Например, рестриктаза EcoPI разрезает нить ДНК между аденином и гуанином, находящимися в следующей последовательности. Г. ААТТ или ТТАА. Г (точка указывает место разрезания ДНК). При этом образуются «липкие концы» — комплементарные друг другу последовательности нуклеотидов — ААТТ и ТТАА, благодаря чему они могут соединяться.
Векторами могут быть плазмиды, бактериофаги, вирусы, космиды. Векторы включают фрагменты ДНК, соответствующие определенному гену, и переносят их в клетку-реципиент.
Плазмиды — мелкие кольцевые молекулы ДНК, присутствующие в клетках бактерий. Они содержат дополнительную генетическую информацию, способны автономно, независимо от ДНК хромосом, реплицироваться; некоторые плазмиды обладают способностью встраиваться в хромосому бактерии и выходить из нее; некоторые могут переходить из одной клетки в другую. В генетической инженерии наиболее широко используются 3 типа плазмид, обозначаемые символами F, Р и Col.
В качестве вектора используют так называемые рекомбинантные (гибридные) плазмиды, в ДНК которых включены гены, выделенные из ДНК прокариот или эукариот. Метод создания рекомбинантных плазмид — векторов был разработан П. Бергом с сотрудниками в 1972 г. Ими была создана рекомбинанрная плазмида, содержащая галактозный оперон Е. coli. В настоящее время известно более 30 систем, позволяющих получать различные векторы. Примером создания рекомбинантных плазмид может быть следующий метод. ДНК плазмиды (например, ПБР 322) и ДНК интересующего исследователя организма обрабатывают ферментом рестриктазой (например, Бам 1), которая образует разрывы ДНК в строго определенных местах с образованием липких концов. Затем оба препарата смешивают, фрагменты ДНК плазмиды и ДНК донора соединяются липкими концами и образуется рекомбинантная молекула ДНК. На соответствующих селективных средах отбирают рекомбинантные плазмиды, содержащие соответствующий ген.
Д. Хелинский с сотрудниками в 1974 г. включил гены, кодирующие синтез аминокислоты триптофана в ДНК плазмиды ColEl, и перенес рекомбинантную плазмиду в клетки кишечной палочки Е. coli. Затем бактерию обработали хлорамфениколом; число копий данной рекомбинантной плазмиды увеличилось до 400—500 на одну клетку, и она стала суперпродуцентом аминокислоты триптофана. Таким путем были созданы штаммы бактерий-суперпродуцентов вакцин к вирусам гепатита В, ящура, гриппа, аденовирусу и др.
В плазмиду могут быть включены природные или синтезированные гены. После проникновения в клетку бактерии рекомбинантная плазмида может функционировать и размножаться автономно либо включаться в ДНК хромосомы бактерии. Таким методом в клетки бактерий были введены гены человека и созданы штаммы бактерий-суперпродуцентов соматостатина, интерферона, брадикинина, гормоны роста человека, быка и других животных, глобин животных и человека (рис. 42).
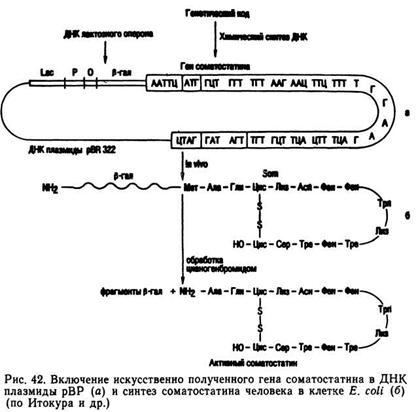
В 1980 г. с помощью плазмиды в клетки Е. coli был введен ген, контролирующий синтез инсулина человека. Для этого из клеток человека была выделена зрелая мрРНК, кодирующая синтез инсулина. С помощью обратной транскриптазы с этой мРНК была получена комплементарная копия — кДНК. Цепочка мРНК была разрушена и с помощью фермента ДНК-полимеразы синтезировали вторую комплементарную нить ДНК. Чтобы синтезированный ген можно было встроить в вектор, к его концу с помощью фермента лигазы были «пришиты» короткие нуклеотидные последовательности — линкеры, которые узнаются рестриктазой Бам 1. Плазмиду и кДНК обрабатывают рестриктазой Бам 1, затем ферментом лигазой и получают рекомбинантную плазмиду, которую вводят в клетку бактерии, приобретающей способность синтезировать про-инсулин.
Вирусы часто используют в качестве векторов, проникающих в животные клетки. Наиболее широко в генетической инженерии используют обезьяний онкогенный вирус ОВ40 (SV40). Он относится к мелким вирусам, его ДНК состоит из 5200 нуклеотидных пар. Геном этого вируса обладает способностью встраиваться в хромосомы клеток млекопитающих. Иногда вирус ОВ40 превращается в вирион, в котором внутри белковой оболочки (капсиды) содержится не ДНК вируса, а ДНК клетки-хозяина. С помощью ОВ40 гены β-цепи гемоглобина мыши и кролика были перенесены в клетки обезьян, где они активно функционировали.
Космиды — векторы, полученные путем объединения небольших фрагментов ДНК бактериофага λ и плазмид. Космиды содержат гены, обеспечивающие их размножение в бактерии, ген устойчивости к антибиотику тетрациклину и особый участок из фага λ под названием «кос», который содержит все вещества, необходимые для упаковки рекомбинантной ДНК в белковую головку фага. Космида состоит из 35— 40 тыс. нуклеотидных пар. С помощью этих векторов гены могут быть перенесены в бактериальные, растительные и животные клетки, культивируемые in vitro.
Большой интерес представляет перенос генов непосредственно в клетки животных, но эта проблема находится пока еще в стадии разработки. В 1982 г. Р. Палмиттер с сотрудниками ввел в мужской пронуклеус оплодотворенных яиц мыши ген гормона роста крысы. Вектором служила рекомбинантная плазмида pMGH, с которой был соединен ген. Ген гормона роста крысы содержит 353 нуклеотидных пары ДНК. Он был введен в виде раствора, содержащего около 600 копий рекомбинантной плазмиды, в 170 яйцеклеток мыши, которые затем были трансплантированы в матки мышей-воспитательниц. Была получена 21 особь, шесть из них проявили гигантизм. В клетках печени гигантских особей содержалось большое количество молекул мРНК, кодирующих синтез гормона роста, а в крови отмечена высокая концентрация этого гормона.
Все увеличивающееся количество данных о расшифровке куклеотидной последовательности в ДНК на участках соответствующих генов обусловило необходимость создания специальных автоматизированных систем для их хранения и обработки. Такие системы созданы у нас в стране, в США и во многих странах Западной Европы.
Генетическая инженерия на уровне хромосом и геномов. Одним из разделов генетической инженерии является разработка методов по экспериментальному переносу из одной клетки в другую целых хромосом. Метафазные хромосомы, выделенные из клетки-донора, могут внедриться в клетку-реципиент путем пикноцитоза. Хромосомы, внедрившиеся в чужую клетку, распадаются на мелкие фрагменты; некоторые из них сохраняются на протяжении нескольких поколений в цитоплазме клетки-реципиента. ДНК, содержащаяся в этих фрагментах, может осуществлять синтез полипептидов. Так, например, в клетки мыши (in vitro) была перенесена 17-я хромосома человека, содержащая гены, контролирующие синтез тимидинкиназы и галактокц-назы. При размножении в мышиных клетках данные гены довольно стойко функционировали. приводит пример использования данного метода в медицине при лечении больных телассемией — тяжелым наследственным заболеванием, обусловленным мутацией генов, кодирующих глобиновые белки —
полипептидные цепи α и β, в результате чего образуются дефектные эритроциты. У больного телассемией берут небольшое количество кроветворных клеток костного мозга, размножают их в культуре вне организма, затем методами генетической инженерии вводят в них полноценные гены, кодирующие глобин, обеспечивающий нормальное развитие эритроцитов. Такие клетки снова вводят в костный мозг того же больного, и они постепенно замещают мутантные патологические эритроциты.
В животноводстве большой интерес представляют методы пересадки клеточных ядер в цитоплазму другого животного — получение цибридов. Например, у мыши извлекали неоплодотворенную яйцеклетку, вводили в нее ядро соматической клетки другого животного и трансплантировали ее в матку самки, гормонально подготовленной к имплантации. Потомство было генетически тождественно той особи, у которой было взято ядро.
Гибридизация соматических клеток. Одной из проблем генетической инженерии является гибридизация соматических клеток, Впервые возможность гибридизации клеток, культивируемых вне организма, установил в 1960 г. Ж. Барский. В 1965 г. г. Харрис обнаружил, что эффективность гибридизации соматических клеток резко повышается при обработке их инактивируемым парагриппозным вирусом Сендай. В настоящее время разработаны и успешно применяются методы, позволяющие добиться слияния клеток различных видов млекопитающих и даже клеток систематически далеких организмов. Например, клетки человека могут сливаться с клетками мыши, крупного рогатого скота, курицы, комара и даже с клетками растений — моркови, табака.
Когда сливаются клетки относительно близких видов, то гибридная клетка может делиться митотически. В процессе деления происходит потеря хромосом одного из видов. Так, в гибридных клетках человек — мышь элиминируются хромосомы человека, что позволяет установить локализацию в них соответствующих генов. Применяя цитогенетический анализ, устанавливают, какая из 23 человеческих хромосом содержится в гибридной клетке. С помощью культивирования их на селективных средах определяют, какие гены в данной хромосоме локализованы. Этим методом в настоящее время локализовано порядка 2000 генов в хромосомах человека.
Получение аллофенных животных. Аллофенными называют химерные организмы, содержащие разные ткани, произошедшие из клеток, полученных от разных родителей. Б. Минтц получил аллофенных мышей путем образования смешанной бластулы из клеток черных и белых мышей (рис. 43).
В последующих опытах соединяли бластомеры животных, различающихся по другим признакам — окраске радужной оболочки, длине ушей и хвоста и др. Для получения аллофенных потомков у беременных мышей, имеющих четко выраженные альтернативные признаки, извлекали эмбрионы на стадии восьми бластомеров и с помощью фермента проназы отделяли бластомеры.
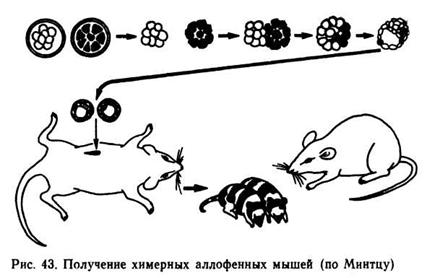
Комбинируя бластомеры от двух (и более) эмбрионов, создали в специальной питательной среде единый комплексный эмбрион, который ввели в матку мыши, гормонально подготовленной к имплантации зародыша. Рождавшиеся мышата представляли собой мозаиков, у них проявлялись признаки всех родительских форм. Методика, разработанная на мышах, в последние годы используется для получения аллофенных овец.
Контрольные вопросы. 1. Докажите, что ДНК принадлежит ведущая роль в наследственности. 2. Каким образом ДНК осуществляет сохранение, передачу и реализацию наследственной информации? 3. Какую роль выполняет каждый фрагмент ДНК, входящий в оперон? 4. Какие свойства присущи генетическому коду? 5. Что такое ген в современном понимании? Каковы функции и свойства генов различных типов? 6. Каким образом осуществляются синтез и выделение генов? Какие векторы вы знаете? 7. Народнохозяйственное значение генетической инженерии.
ГЛАВА 9. ГЕНЕТИЧЕСКИЕ ОСНОВЫ
ИНДИВИДУАЛЬНОГО РАЗВИТИЯ
Процесс индивидуального развития особи — онтогенез — начинается с акта оплодотворения и заканчивается смертью. В процессе онтогенеза реализуется наследственная информация, присущая генотипу данной особи. Она определяет время, место и последовательность развития органов и признаков. Онтогенез запрограммирован в генотипе особи, но осуществляется в конкретных условиях внешней среды, определяющей характер и возможности реализации наследственной информации. При изучении закономерностей индивидуального развития животного или растения большое внимание уделяется процессу формирования органов — органогенезу. Постепенное становление формы и функции каждого органа у данной особи в процессе ее развития определяет морфогенез.
При изучении онтогенеза главной задачей является раскрытие закономерностей конкретной реализации гена: каким образом ген, локализованный в молекуле ДНК, контролирует развитие специфического признака, характерного для вида, породы и отдельной особи.
У прокариот путь от гена к признаку относительно простой; ген контролирует синтез фермента и его активность регулируется процессами, протекающими непосредственно в клетке. Мы можем судить о генотипе данного штамма бактерии Е. coli по его способности синтезировать фермент, аминокислоту, антибиотик или другое вещество или гидролизовать питательный субстрат, на котором он размножается. Благодаря механизму регуляции генетического кода бактерии обеспечивают своевременную активность генов, синтезирующих ферменты, необходимые клетке в данный период ее жизнедеятельности. Следовательно, у прокариот четко прослеживается связь между геном и признаком: ген → фермент → признак.
Значительно сложнее этот процесс осуществляется у высших многоклеточных сложноорганизованных особей. Каждый признак у них контролируется, как правило, многими генами, формируется в онтогенезе под влиянием многих ферментов, во взаимодействии с другими органами и тканями. Например, окраска меха у норок контролируется более чем 20 генами, окраска шерсти у крупного рогатого скота зависит от различного сочетания 10 генов, цвет глаз у дрозофилы зависит от 20 генов. Каждый ген в генотипе особи представлен двумя аллелями, в результате перекомбинации которых может осуществляться самое разнообразное проявление признака. Важное значение для характера проявления признака имеет и возможное взаимодействие генов.
Существенное влияние на характер развития органа и проявления признака могут оказывать условия внешней среды. У животных эмбриональное развитие протекает в организме матери, поэтому на признаки, формирующиеся до рождения, внешняя среда оказывает относительно слабое влияние, опосредованно через организм матери. Они развиваются главным образом под контролем генотипов родительских форм, и после рождения почти не изменяются. К таким признакам в первую очередь относят морфологические особенности строения каждого органа, а также группы крови, типы гемоглобина и др.
В постэмбриональный период условия внешней среды оказывают существенное влияние на характер проявления главным образом количественных признаков, определяющих продуктивность животного. Очень показательны в этом отношении однояйцовые близнецы. Идентичность их генотипов четко проявляется в распределении окраски шерсти, форме рогов, строении конечностей, но форма тела, масса, продуктивность значительно изменяются под влиянием скудного или, наоборот, обильного кормления
Биогенетический закон онтогенеза. Онтогенез имеет генетическую предопределенность развития животных данного класса и свидетельствует об общности их происхождения. Общность происхождения генетически разных форм отражена в чертах их развития и порядке смены этапов и фаз, в появлении у систематически разных групп ряда черт, характерных для их предковых форм.
Онтогенез каждой особи подчиняется биогенетическому закону Мюллера-Геккеля: сходство эмбриональных черт развития отражает степень родства разных форм в силу общности их происхождения. Процесс дробления зиготы у всех многоклеточных животных организмов проходит начальные стадии эмбриогенеза — бластулу и гаструлу. Для позвоночных характерно прохождение стадии, на которой у наземных форм, дышащих легкими, образуются жаберные дуги, как и у рыб.
В филогенетических рядах характерен параллелизм гистологической структуры и функций некоторых тканей — нервной, мышечной, эпителиальной, соединительной. Наличие этой закономерности, установленной русским ученым (1886—1945), отражает общность генетической детерминации клеток в весьма давно и далеко разошедшихся классах, например у млекопитающих и у насекомых.
Вместе с тем необходимо учитывать и различия в онтогенезе у разных форм, которые накладываются на общий характер развития, что свидетельствует о специфике генетической информации. Так, например, в яйцах рыб и птиц содержатся большие запасы желтка, что сказывается на их эмбриогенезе.
Неравномерность и неодновременность процессов роста и дифференцировки. Рост может осуществляться либо за счет деления клеток, либо за счет их растяжения. В период роста органов и тканей преобладают процессы, обеспечивающие митотическую активность клеток. В этот период наиболее активно функционируют гены, контролирующие синтез ферментов, обеспечивающих все периоды и фазы митотического цикла.
В эмбриональной ткани животных клетки относительно одинаковы по форме и составу белков. Позже они дифференцируются, при этом эмбриональная клетка превращается в клетку с различной специализацией. Такое проявление различий между клетками называют дифференцировкой, а клетки — дифференцированными. В период дифференцировки активно функционируют гены, контролирующие синтез специфических белков, необходимых для формообразовательных процессов и интеграции специализированных клеток.
Необратимость и обратимость процесса дифференцировки соматических клеток и тканей. Зигота содержит полный набор генов и всю генетическую информацию данного вида, породы и особи; она тотипотентна (омнипотентна), то есть обладает всеми возможностями развития и формирования органов и признаков взрослой особи. Дифференцировка клеток не приводит к потере ими имеющихся возможностей. Генетическая информация, содержащаяся в молекулах ДНК, одинакова в любой соматической клетке, то есть каждая соматическая клетка потенциально способна дать начало новому организму. Тотипотентность соматических клеток характерна для растений. Из одиночных клеток, выделенных из дифференцированных тканей любого органа, можно в пробирочной культуре получить целое растение, идентичное исходному. Такие растения получают из корнеплодов сахарной свеклы и моркови, из клетки листа бегонии и многих других культур. У животных тотипотентность клеток сохраняется только на ранних этапах онтогенеза. Р. Бриггс и Т. Кинг (1952), Дж. Гёрдон (1964) выделяли ядра из мышечных клеток или из клеток кишечного эпителия головастиков шпорцевой лягушки и пересаживали их в безъядерные активированные яйцеклетки. Из некоторых яйцеклеток с пересаженным ядром соматической клетки развивались нормальные головастики и взрослые особи (рис. 44).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
