
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У кур зафиксировано большое число наследственных аномалий, имеющих рецессивный, доминантный и сцепленный с полом тип наследования. Рецессивные аномалии: неспособность к вылуплению; укорочение верхней челюсти и клюва; дефект маховых перьев; уродства позвоночника и таза; уменьшение глазного яблока и гибель сразу после вывода, укорочение и утолщение конечностей; многопалость; отсутствие нижней челюсти и мозговая грыжа; бескрылость и отсутствие легких, почек и воздушных мешков; карликовость; запрокидывание головы и дрожание; гипоплазия конечностей и др. Среди доминантных аномалий кур обнаружены коротконогость (летальность для гомозигот); врожденное дрожание; отсутствие оперения; атрезия яйцевода и др. Выявлены аномалии, обусловленные сцепленным с полом наследованием и приводящие к отсутствию оперения у курочек, внезапной гибели курочек в возрасте до 123 дней. Обнаружены летальная черная окраска; «трясучка», поражающая молодняк 2—5-месячного возраста; волокнистый пух, приводящий к гибели в возрасте 14 дней; одышка, пароксизм (угнетение роста) и др.
Каждому специалисту необходимо тщательно описывать все проявляющиеся в стаде аномалии или патологические признаки, регистрировать частоты их возникновения и выявлять родственную связь между аномальными животными и их предками, сибсами, полусибсами и др. Необходимо планировать отбор и подбор пар таким образом, чтобы стало невозможным дальнейшее распространение летальных или других нежелательных генов в породе.
Наследственность основных массовых болезней и проблема селекции на резистентность. Более важное значение для практики имеет наследственная устойчивость (резистентность) организма к ряду заболеваний, затрагивающих не единичных особей в стаде или породе, а распространяющихся на значительное поголовье и наносящих большой экономический ущерб. Наиболее опасными по своему патологическому, экономическому эффекту и трудностям в их ликвидации обычными ветеринарными приемами являются инфекционные и инвазионные болезни (бруцеллез, туберкулез, лейкоз, маститы, рожа, пироплазмоз, пуллороз кур, птичий тиф и др.).
Традиционные ветеринарные методы лечения, лежащие в основе очищения стад от некоторых заболеваний, дают эффект в основном в тех группах животных, которых подвергали прививкам и у них выработался пассивный иммунитет. Для последующих поколений вновь потребуются такие же мероприятия. При некоторых заболеваниях вынужденно применяются массовый убой и ликвидация животных, особенно если против распространяющейся болезни не разработаны ни профилактические, ни лечебные мероприятия. Вынужденный убой животных — это крайняя мера, поэтому необходимо вести селекцию на создание стойкой резйстентностй животных и закреплять ее в ряде поколений.
Устойчивость животных к указанным заболеваниям имеет полигенный тип наследования, то есть обусловлена действием многих генов. Выявление генетического детерминирования некоторых заболеваний создает основу для осуществления селекции на резистентность. У животных с большим интервалом Между поколениями (у крупного рогатого скота интервал Составляет около пяти лет) темп селекции на резистентность будет медленнее, чем у животных с малым интервалом между поколениями (птица), характеризуемых высоким коэффициентом размножения Селекция на резистентность усложняется и тем, что отбор ведут одновременно по нескольким признакам.
На формирование резистентности и эффект селекции по ее показателям влияют условия внешней среды (уровень и тип кормления, параметры микроклимата и др.). Эти факторы могут неблагоприятно отразиться на здоровье животных и тем самым затормозить селекцию на резистентность.
При селекции на резистентность пользуются двумя методами. Один из них основан на искусственном заражении животных патогенными микроорганизмами. На фоне такого заражения часть животных гибнет или их выбраковывают, а часть не реагирует на заражение, что обусловлено индивидуальной наследственной резистентностью. Эту группу животных используют для дальнейшего размножения и селекции на резистентность потомства последующих генераций. Метод не может быть применен в производственных условиях.
Другой метод основан на проведении генетического анализа семейств, что дает возможность выявить более и менее резистентных животных и осуществить селекцию в нужном направлении.
Определенные затруднения в селекции на закрепление резистентности к инфекционным болезням возникают в связи со способностью патогенных микроорганизмов проявлять большую изменчивость, при которой за короткие отрезки времени один и тот же вид бактерий или вирусов изменяет наследственность. В результате этого животные, резистентные к одному штамму, оказываются восприимчивыми к вновь возникшему штамму микроорганизма. Селекцию на резистентность животных усложняет и родственное спаривание. Инбридинг приводит к повышению гомозиготности стад и пород, часто вызывает инбредную депрессию, снижает резистентность инбредного потомства, увеличивает распространение в популяции нежелательных рецессивных генов и гомозиготных (часто летальных) генотипов.
Несмотря на трудности в селекции на резистентность, получены обнадеживающие результаты по созданию резистентных групп свиней, крупного рогатого скота и птицы.
Использование селекционных методов создания и выведения резистентных популяций сельскохозяйственных животных осуществляется в нашей стране ведущими научно-исследовательскими коллективами (ВНИРГЖ, ВИЖ, ВНИИТИП, МВА и др.) При этом основное внимание направлено на создание резистентных популяций животных разных видов к таким распространенным заболеваниям, как маститы, лейкозы сельскохозяйственных животных. В птицеводстве проводится работа по борьбе с болезнями Марека, туберкулезом, бруцеллезом, пуллорозом, кокцидиозом, колибактериозом птицы и др.
Наряду с работами, практически доказавшими возможность выведения резистентных групп животных, многие исследования, направленные на разработку проблемы повышения естественной резистентности, носят поисковый и экспериментальный характер. Вместе с тем они позволяют накапливать данные, подтверждающие генетическую обусловленность индивидуальной и групповой естественной резистентности и разрабатывать селекционно-генетические методы предупреждения и снижения заболеваемости животных.
Существенный урон наносит распространение маститов у крупного рогатого скота в острой или субклинической форме. Повышение заболеваемости маститом наблюдается в зимне-стойловый период и несколько уменьшается в летне-пастбищный, когда благоприятное действие оказывает ультрафиолетовое естественное облучение, вызывающее гибель микрофлоры. Увеличение возраста коров сопровождается распространением острой и субклинической форм мастита, что связано с общим возрастным ослаблением защитных средств, а также с ослаблением сфинктера канала соска, способствующим доступу микрофлоры в сосок,
При анализе генеалогической структуры стада в исследованиях ряда научных коллективов (ВИЖ, ВНИРГЖ, МВА и др.) была выявлена связь маститоустойчивости у некоторых генетико-селекционных групп коров. Отмечено, что в некоторых семействах и линиях это заболевание отсутствует, в то время как в том же стаде другие группы проявляют высокую заболеваемость (до 30—50% от числа всех животных группы). При инбридинге с родительских пар снижайся потомства.
Для снижения заболевания маститом в настоящее время осуществляют селекцию на правильную форму вымени, устойчивость нервной системы коров, при которой наблюдается меньше «срывов» в молокоотдаче. Вместе с тем необходимо проводить отбор животных, обладающих высоким уровнем наследственно обусловленной естественной резистентности тканей молочной железы к патогенным микроорганизмам и способности клеток синтезировать защитные вещества, входящие в состав крови, молока, слизи, в частности такой фермент, как лизоцим,
Так, в работах и др. (1977) была обнаружена различная степень устойчивости коров голландской и шведской пород к субклиническому маститу. У голландской породы количество больных коров достигало 17%, а у шведских —25%. Выявлены более резистентные семейства и линии. Отмечено, что в группах здоровых коров частота аллеля гена βLgA в белке молока выше, чем в группе больных. В тех семьях, где заболевание маститом не отмечено частота этого аллеля составляла 0,4865 и определено большое число животных, гетерозиготных по локусу βLg (59,5%).
В экспериментах с айрширским скотом (Меркурьева, Скрипниченко, Беляева, 1980) установлено, что по такому показателю естественной резистентности, как активность лизоцима в крови, молоке и молозиве коров, дочери разных быков существенно различались Различия между дочерьми разных быков были выявлены и по уровню бактерицидной активности. Заболевание коров маститом варьирует у животных разных семейств
Установлена достоверная разница в активности лизоцима сыворотки крови коров, имеющих разные генотипы по локусам трансферрина, амилазы и церулоплазмина сыворотки крови, что создает возможность отбора желательных генотипов (TfAA, TfAE, АтСС, CpBB) для повышения естественной резистентности.
У коров со здоровым выменем активность лизоцима была ниже как в молозиве, так и в молоке. У коров со скрытыми (субклиническими) формами мастита эти показатели были достоверно в два раза выше, что являлось результатом мобилизации защитных механизмов против маститной инфекции. Таким образом, тестирование животных по уровню лизоцимной активности создает перспективу включения в селекционный процесс новых показателей с целью повышения естественной резистентности
Большой экономический ущерб скотоводству наносит лейкоз, поэтому в последние годы многие исследования направлены на выявление наследственной обусловленности этого заболевания. Различают «вертикальный» тип распространения лейкоза, когда он передается из поколения в поколение, и «горизонтальный» тип, когда он распространяется между хозяйствами в результате переноса возбудителя,
Существует ряд теорий этиологии лейкоза и его генетического детерминирования, но достаточной ясности в этом вопросе пока нет, Вирусная теория происхождения лейкоза исходит из признания наличия онкогенного возбудителя. Вирус может находиться в латентном состоянии, а при определенных условиях переходит в активную форму. Он может передаваться от матери к плоду через плаценту, через молозиво и приводит к картине «семейного» и «врожденного» лейкоза. Вместе с тем в этиологии и распространении лейкоза имеет большое значение наследственность животного. Многими исследованиями установлено, что можно выделить лейкозоустойчивых и, наоборот, подверженных этому заболеванию животных.
В 1968 г. сформулирована вирусно-генетическая теория возникновения и распространения лейкоза. Считают, что восприимчивость к лейкозу контролируется доминантными, а устойчивость к нему — рецессивными аллелями аутосомных хромосом животного. Между онкогенными вирусами и клетками организма животного, подверженного заболеванию, существует определенное взаимодействие. Размножение вируса может происходить только при внедрении его в клетку животного, в результате чего наступает процесс репликации РНК вируса. При этом вирус вызывает большие изменения в морфологии и обменных процессах зараженных клеток.
Исследованиями, проведенными на большом поголовье скота красной степной породы, был подтвержден наследственный характер лейкоза, который прослеживался в нескольких поколениях. Ею сформулирована гипотеза о том, что в основе заболевания лейкозом лежит провирус (V), ДНК которого включается в геном клетки крупного рогатого скота. Активность провируса зависит от наличия в генотипе животного доминантного гена-репрессора (R) или его рецессивного аллеля r. Если в генотипе клеток рецессивный ген будет находиться в гомозиготном состоянии (rr), то провирус становится активным, что приводит к заболеванию животного лейкозом. Следовательно, наличие провируса V и аллеля г создает состояние предрасположенности к лейкозу и при генотипе клеток животного V— rr проявляется заболевание.
Наследственная обусловленность резистентности животных к лейкозу не вызывает сомнения и подтверждена обширными материалами многих исследователей (Емельянов и сотр., 1966; Визнер, 1967; Лактионов, 1968; Эрнст, Цалитис, 1973) Было доказано, что устойчивость к лейкозу обусловлена полигенным (полимерным) типом наследования (Петухов, Карликов, 1981). Коэффициент наследуемости (h2) резистентности к лейкозу значительно колеблется (от 0,10 до 0,33 и более) у животных разных стад и разного происхождения.
А. С Емельянов показал, что заболевшие лейкозом коровы черно-пестрой породы были дочерьми, внучками и правнучками быка Прибоя и его сына Таинственного Доказано наследование лейкоза у бурого латвийского скота в зависимости от принадлежности животных к семействам и линиям.
В процессе исследований, проводимых академиком В. П. Шишковым с сотр. (1983), выдвинута вирусно-иммуногенетическая теория этиологии, патогенеза и профилактики лейкозов и других опухолевых заболеваний. Показано, что вирусы, вызывающие онкогенные заболевания, интегрированы с геномом кроветворных клеток. Развитие лейкоза в организме связано с иммунобиологическим состоянием животного и его генетической предрасположенностью к заболеванию. Противолейкозный иммунитет связан с состоянием Т - и В-лимфоцитов, макрофагов и неспецифических факторов резистентности организма.
Установлена связь заболевания лейкозом с полиморфными системами крови животных. Так, по данным и др., оказалось, что коровы, гомозиготные по аллелю TfA (генотип TfAA), проявляли более высокую восприимчивость к лейкозу и около 36% животных с таким генотипом болели лейкозом, а при гетерозиготных генотипах (TfAD, TfED) насчитывалось только 20—27% больных животных.
По данным , , Д. В, Карликова, выявлены существенные различия между носителями некоторых аллелей и генотипов B-системы групп крови быков и заболеваемостью лейкозом их дочерей. Так, в потомстве быков, маркированных аллелем В1р´ установлены очень редкие случаи заболевания лейкозом, а дочери быков, имевшие альтернативный аллель, часто болели.
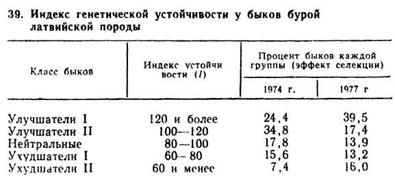
Для селекционных целей разработан популяционный коэффициент (I) — индекс генетической устойчивости. При обследовании скота бурой латвийской породы были получены данные, представленные в табл. 39.
Данные табл. 39 свидетельствуют о том, что чем больше индекс устойчивости, тем выше резистентность. Ежегодно такая оценка быков дает возможность выявить около 25% быков-улучшателей по генетической устойчивости к лейкозу. Эффективность отбора по устойчивости к лейкозу в семействах значительно ниже, чем по линиям. В экспериментальном хозяйстве «Сигулда» Латвийского научно-исследовательского института животноводства и ветеринарии этот эффект составил около 3% на одно поколение.
Выявлены породные особенности восприимчивости и устойчивости крупного рогатого скота к лейкозам Повышенная заболеваемость отмечена у черно-пестрого фризского скота, у канадских голштинов (Roberts, 1980), у финских айрширов (Neuvonen и др., 1986). Исследователями США (Haus и др., 1977) выяснено, что поражаемость бычьим лейкозным вирусом (БЛ) у мясного скота составила 2,6%, у молочного — 28,2% (особенно у голштино-фризов).
По данным ВНИИПлема (1983), распространение лейкоза в нашей стране по разным породам варьирует от 0,4—0,8% для симментальской, швицкой, холмогорской пород до 10,3-22% для черно-пестрой и голштино-фризской пород. По данным этих авторов, оказалось, что среди коров, заболевших лейкозом, большая часть получена с использованием инбридинга Вирус лейкоза может передаваться потомству в эмбриональный период от больной матери и через молозиво
Выяснено, что у животных, гомозиготных по группам крови, восприимчивость к лейкозу меньше у больных коров эстонской породы чаше встречаются эритроцитрные антигены Т, Е, С, М; у черно-пестрой -I, P, Т, О, ОА, КZ, X и С.
На основании обобщения работ по изучению лейкоза ВНИИРГЖ выпустил для использования в практика «Рекомендации по селекции крупного рогатого скота на устойчивость к лейкозу» (Л. 1986), где приведены методы оценки быков на лейкозоустойчивость и схема селекционной работы
Для оценки быков на лейкозоустойчивость сравнивают процент заболе ваемости по стаду (С) со средним процентом заболевших коров-сверстниц (В). Формула лейкозоустойчивости быков (О) по показателям дочерей такова
![]()
где Д — число эффективных дочерей, которое определяется по формуле
Дэ=nдочерей×nсверстиц/nдочерей+nсверстниц; Д - заболеваемость дочерей, %;
В — заболеваемость сверстниц, %; 39 — постоянный коэффициент при наследуемости h2=0,l
Если известна заболеваемость среди матерей дочерей оцениваемого быка, то в формуле учитывают этот показатель
![]()
где М — заболеваемость матерей дочерей оцениваемого быка,%; Вм — заболеваемость матерей сверстниц, %; 0,3 — регрессий дочерей на матерей по заболеваемости.
Если требуется сравнить быков, содержащихся в разных условиях, то применяют индекс генетической устойчивости (И):
![]()
В рекомендациях дается схема проведения селекции крупного рогатого скота на устойчивость к лейкозам (рис. 63).
Быков считают лейкозоустойчивьми, если ни одна из их дочерей не болела лейкозом до окончания третьей лактации Условно устойчивыми считают быков, у которых процент лейкозных дочерей меньше, чем процент больных сверстниц в стаде. Восприимчивыми быками будут те, у которых процент заболевших дочерей равен или выше процента больных среди сверстниц стада.
Комплекс селекционных мероприятий предусматривает проведение оценки быков по состоянию здоровья дочерей, недопущение внутрилинейного подбора, а группах скота, неблагополучных по лейкозу.
Продолжительные исследования проведены (1963—1986) по изучению генетики устойчивости животных к лейкозу и другим заразным болезням у ряда пород молочного скота из разных зон нашей страны. Популяционный анализ показал, что коэффициенты наследуемости устойчивости составляли по лейкозу 0,3, по бруцеллезу 0,194, туберкулезу 0,10, маститу 0,10, болезням конечностей 0,13.
Было установлено, что от быков, у которых отмечен высокий процент дочерей, болевших лейкозом в данном хозяйстве, эти особенности проявлялись и в других хозяйствах, что подтверждает оценку быков как носителей наследственно обусловленной лейкозоподверженности. Генетико-селекционный анализ выявляет лейкозоустойчивость потомства, принадлежащего к разным линиям и семействам. Такие свойства отдельных групп прослежены в нескольких поколениях потомков. Целесообразно проводить селекцию на лейкозоустойчивость в стадах и в породе, используя положительно оцененные группы (семейства, линии, отдельные производители) быков и коров.
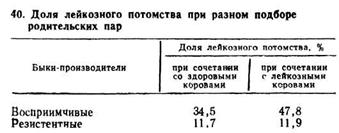
По данным , имеются различия при подборе резистентных (или восприимчивых) быков к здоровым или лейкозным коровам (табл. 40).
Заболеваемость крупного рогатого скота туберкулезом и бруцеллезом также имеет наследственную обусловленность. Выделяются как семейства коров, так и потомство некоторых быков, проявляющих более высокую (или более низкую) заболеваемость такими болезнями. По данным , для туберкулеза в среднем по популяции h2 = 0,10, а по разным линиям его величина колебалась от 0,01 до 0,492. Коэффициент наследуемости бруцеллеза в среднем составил 0,194.
Для повышения резистентности к этим заболеваниям недостаточно проводить массовую селекцию или выбраковывать больных животных. В современной селекции необходимо проводить планомерный подбор пар с учетом использования резистентных животных, устранять из подбора восприимчивых и проводить индивидуальную оценку производителей и маток по их личной устойчивости к источнику болезни и по устойчивости их потомков. При этом необходимо вести такую оценку не по одному заболеванию, а по их комплексу, имея в виду, что связь между заболеваниями существенна. Она выражается коэффициентами корреляции на уровне r = 0,383 по семействам, а по отцам в дочернем поколении составляет r=0,340.
Следовательно, для осуществления селекционной работы на повышение резистентности к массовым заболеваниям необходимо вести учет и регистрацию в племенных индивидуальных документах заболеваний, подвергая затем эти данные генетическому и популяционному анализу. В системе крупномасштабной селекции ведение записей и анализ динамики по поколениям и календарным отрезкам должны быть обязательным элементом работы зоотехника-селекционера и ветеринарного врача,
Многочисленные исследований проведены по изучению резиетентноссти у птицы. В числе ранних работ по получению резистентных групп следует назвать Ф. Хатта (1954), который путем отбора получил устойчивую к пуллорозу линию кур породы белый леггорн. При этом было применено экспериментальное заражение птицы. В процессе селекции в зараженной группе птицы в результате отбора сохранность достигла 70%, а в контрольной — только 28%, Скрещивание резистентной птицы с контрольной показало, что в потомстве первого поколений наблюдалось наследование резистентности, что указывает на доминантный тип наследований этого признака.
Эффект успешной селекции был получен этим же исследователем в результате 15-летней работы по выведению птицы» устойчивой к лейкозу. Была получена лейкозоустойчивая линия, в которой отход на протяжении от 40-До 600-дневного возраста составлял 5%, а в лейкозочувствительных линиях он достигал 60%.
Ряд опытов был посвящен селекции птицы, резистентной и чувствительной к заражению вирусом саркомы Рауса (ВСР). Отобранные группы птицы, проявившие устойчивость к лейкозу, послужили основой для создания новых резистентных линий. Отбором для размножения более резистентных к саркоме кур удалось снизить количество больной птицы с 70 до 12,6%, В линии, отселекционированной на повышенную чувствительность к лейкозу, доля больной птицы в Четвертом поколении достигла 78%
Выявлена разная степень породной устойчивости птицы (леггорн, полтавские глинистые куры), их помесей и гибридов. У помесей (полтавская глинистая × род-айланд) индекс резистентности составил 51—-59%, а у леггорнов — 13 — 14%. Наследуемость резистентности была значительной (h2 =0,33 — 0,59), причем коэффициент наследуемости по отцу был выше, чем по матерям
Исследований, проведенные в МВА (Бессарабов, 1983), показали эффективность повышений генетической устойчивости сельскохозяйственной птицы. Разработана методика отбора кур на естественную резистеитность против пуллороза, колибактериоза, стафилококкоза путем заражения птицы оттитрованным материалом, что позволило дифференцировать и оценивать кур, устойчиво передающих резистентность потомству.
Влияние генетических различий на показатели естественной резистентности получило подтверждение в опыте (Меркурьева, Саладдин, 1981) при сравнений уровня иммунологических показателей сыворотки крови цыплят кросса «Беларусь-9» и его исходных линий. Гибридная птица имела более высокий уровень лизоцима, бактерицидной активности и бета-лизина по сравнению с исходными линиями, причем последние также различались между собой по имунным показателям. Были также проведены опыты с птицей трех различных кроссов и типа продуктивности: бройлеры, яичные кроссы «Хайсекс белый» и «Хайсекс коричневый». Изучены показатели активности лизоцима, бактерицидной активности, бета-лизина, количество эритроцитов, величина гематокрита. Уровень лизоцима в белке яиц кросса «Хайсекс коричневый» был выше, чем в других кроссах, что способствует повышению выводимости и резистентности цыплят этого кросса (Меркурьева, Айтман, 1987). Доля влияния генетических различий на комплекс факторов резистентности белка яиц, крови, эритроцитов была достоверна и отражала влияние селекции на резистентность
Обработка яиц перед инкубацией парами супермутагена нитрозодиметилмочевины (НДММ) в микродозах 1 10-6 г/яйцо дала возможность повысить активность лизоцима в белке на 10%, а в сыворотке крови суточных и месячных цыплят, полученных из обработанных яиц, на 15% по сравнению с уровнем у птицы контрольной группы. При заражении 45-дневной птицы кросса «Беларусь-9» сублетальной дозой культуры микоплазмы установлено, что уровень лизоцима, бактерицидной активности и бета-лизина был выше у птицы, полученной из яиц, обработанных парами НДММ, как в группе незараженных, так и в группе зараженных цыплят. Более высокая естественная резистентность птицы, полученной из обработанных яиц, подтверждалась более высокой их сохранностью при выращивании по сравнению с контрольной птицей (Красота» Шангин-Березовский, Молоскин, Саладдин, 1981).
Исследования гельминтологов показали, что наблюдается наследственная устойчивость кур к аскаридозу, что связано с наличием определенных генотипов по полиморфным системам некоторых ферментов. Наиболее подвержена аскаридозу птица, характеризующаяся сочетанием следующих генотипов по этим же локусам: PpFF, EsBB, CaFF, HbBB, то есть имеющих гомозиготную форму (Селихова, 1983).
Эффективные результаты получены при селекции рыб на резистентность Целью работы было создание групп карпа, резистентных к краснухе (Илясов, Кирпичников, Шарт, 1978). Для этого использовали ропшинскую группу (Р), украинско-ропшинских помесей (УР) и местного краснодарского (М) карпа. Селекцию проводили на фоне естественного и искусственного заражения рыб и жесткого отбора в течение 4—5 поколений. Число выживших рыб, содержавшихся в условиях очень уплотненной посадки, вызвавшей вспышку краснухи, указывает на высокий эффект селекции Резистентность рыбы повышалась следующим образом.

Помесная рыба отличалась эффектом гетерозиса Карпы, имевшие генотип по трансферрину TfAB и эстеразе EsFF, характеризовались повышенной устойчивостью к краснухе.
Реализация генетически обусловленной резистентности может усиливаться различными методами кормления, содержания и биологической стимуляцией. Как показали исследования, селекцией можно повышать не только резистентность животных к болезням, но и устойчивость к неблагоприятньм факторам среды, Устойчивость к высокой температуре у крупного рогатого скота связана с понижением функции щитовидной железы. Так, бирманский зебувидный скот, у которого функция щитовидной железы понижена, более устойчив к высоким температурам; у герефордской породы функция этой железы ярко выражена и скот плохо переносит жару
А, Н. Соколовой (ВНИИРГЖ, 1960—1986) путем длительной селекции была создана линия кур, приспособленных к низким температурам окружающей среды при сохранении высокой яйценоскости. В течение девяти поколений цыплят с первых дней жизни выращивали при температуре 12 — 16°С, что формировало конституционально крепкую птицу с хорошей приспособленностью и повышенной терморегуляцией.
Птица проявляла комплексную устойчивость к аскаридозу, кокцидиозу, болезни Марека и карциноме. Эти качества резистентности сочетались со скороспелостью; яйценоскость за первые 73,5 нед составляла в среднем 244 яйца при рекорде 303—310 яиц, что соответствует мировым стандартам. Была получена и линия кур русской белой породы, которая обладала повышенной резистентностью к онковирусам RPV. При разведении на протяжении 18 поколений птица сохраняла свои свойства на фоне внутрилинейного подбора при коэффициенте инбридинга от 30 до 70%.
(1977) изучал приспособленность животных к неблагоприятным условиям среды. Была высказана гипотеза, что в процессе доместикации у животных ослаблена общая адаптационная пластичность и снижена неспецифическая, то есть естественная, резистентность. Выяснено, что важную роль в эколого-генетических свойствах животных играет биохимический полиморфизм белковых систем крови, в частности полиморфные типы гемоглобина. К экстремальным условиям высокогорья лучше приспособлены животные с гетерозиготным генотипом гемоглобина (НbАВ) и с генотипами, обеспечивающими высокое содержание в крови концентрации калия,
Немалое значение для формирования естественной резистентности имеет кормление. Выла установлена (Красота, Дронин, 1980) связь между особенностями рациона (брикетный, корнеплодный и концентратный типы кормления) телок костромской породы и уровнем показателей их естественной резистентности. У телок, получавших корм в виде брикетов, бактерицидная активность была достоверно выше в период от одного до 18-месячного возраста. Показатели иммунологической реактивности и естественного иммунитета (количество лейкоцитов, лизоцимная и бактерицидная активность и фагоцитарная реакция) свидетельствовали о повышении стабильности гуморальных и клеточных факторов, обеспечивающих устойчивость животных к микробным инфекциям. Это сопровождалось хорошей минерализацией костяка, более высокой репродуктивной способностью телок и более высокими экономическими показателями. Самые благоприятные результаты получены при выращивании телок на брикетах клеверотимофеечной резки.
Работами и (1981—1986) было показано, что подкормка телят черно-пестрой породы балансирующими витаминно-минеральными добавками не только улучшала развитие молодняка, но и повышала уровень и стабильность комплекса факторов естественной резистентности.
Обобщение материалов, полученных в исследованиях на различных видах и породах животных, дает возможность сделать вывод, что в задачу генетики входит разработка методов, позволяющих выявлять наследственную патологию (аномалии, уродства, болезни), устанавливать показатели, характеризующие степень резистентности животных или их предрасположенность к заболеваниям. Имеющиеся в этом направлении достижения науки и практики подтверждают возможность осуществления селекционно-генетического оздоровления отдельных групп и популяций животных.
Контрольные вопросы. 1. Дайте характеристику неспецифической и специфической защитной функции организмов и укажите, какие органы несут ведущую роль в создании этих функций. 2 Клеточные факторы защиты, роль клеток белой крови, их развитие в онтогенезе 3. Иммуноглобулины, их классы, место синтеза и основные особенности в структуре молекулы, 4. Наследственные болезни у основных видов сельскохозяйственных животных. 5. Методы выявления наследственных аномалий и болезней. 6. Использование индекса генетической устойчивости в селекции скота. 7. Генетический полиморфизм белковых систем у птицы и связь с ними резистентности.
ГЛАВА 16. ИММУНОГЕНЕТИЧЕСКИЙ И БИОХИМИЧЕСКИЙ БЕЛКОВЫЙ ПОЛИМОРФИЗМ
Генетический полиморфизм отражает важнейшие особенности внутривидовой и межвидовой изменчивости, обусловленной наследственностью. Под генетическим полиморфизмом понимают наличие в популяции одновременно нескольких аллельных состояний гена конкретного локуса, определяющих формирование разных фенотипов данного признака. Термин «полиморфизм» введен Е. Фордом в 1945 г. применительно к различным признакам, обусловленным наследственностью.
Наличие в локусе нескольких аллельных состояний гена увеличивает генетическую изменчивость в популяции и сопровождается образованием гетерозиготных генотипов, которые благоприятствуют выживанию гетерозиготных особей Гетерозиготность локуса делает возможным подавление доминантным аллелем А вредных рецессивных аллелей а (А>а) или создание преимущества для организма за счет сверхдоминирования, когда генотип Аа оказывает большее положительное влияние, чем генотип АА (Аа>АА).
Гетерозиготность локуса может проявляться как в структурах иммунных систем антигенов, повышая защитные функции организма, так и в виде биохимического полиморфизма, когда разнообразие белковых или ферментных веществ, обусловленных разными аллелями данного локуса, создает в организме возможности более гибкого взаимодействия со средой
В практике селекции важное значение имеет обнаружение проявляющих полиморфизм наследственно обусловленных признаков, которые либо сами являются предметом селекции, либо используются в косвенной селекции в качестве генетических маркеров хозяйственно полезных признаков, на совершенствование которых должна быть направлена селекция
Наследственно детерминированные биологические системы, такие как иммуногенетические образования в виде групп крови, а также генетически обусловленные полиморфные биохимические вещества (белки и ферменты крови, молока и других тканей организма), подтверждают наличие генетического полиморфизма. Их применение основано на том, что группы крови и полиморфные системы белков не изменяются в процессе онтогенеза и являются пожизненной генетической характеристикой каждой особи.
Иммуногенетика. Новое направление в иммунологии начало формироваться с открытием К. Ландштайнером в 1900 г специфических реакций эритроцитов крови, происходящих при переливании крови у человека. Было показано, что существует определенная система эритроцитарных групп, которые были названы группами крови. К настоящему времени у человека зарегистрировано 14 эритроцитарных систем групп крови. Генетику групп крови начали исследовать Дунгер и Гиршфельд (1910 г.), затем это было дополнено и получило объяснение в работах Бернштейна (1924 г.)
В 1947 г новое направление в биологии было названо иммуногенетикой. В основе ее объединены иммунологические и генетические методы, выявляющие особенности реакции между эритроцитарными антигенами и антителами.
Иммуногенетические исследования применительно к животным были начаты в работах американских исследователей Оуэна, Стормонта и Ирвина (1944), затем Неймана и Серенсена (1956), Ренделя (1958). С 1957 г создана школа иммуногенетиков в Чехословакии (Матоушек) В 60-х годах развернуты исследования в Венгрии (Ромвари и др., 1961), в ГДР (Буш, 1963), Японии (Хосода, 1965) и в других странах
В нашей стране исследования в области иммуногенетики у сельскохозяйственных животных были начаты в 60-е годы. В настоящее время работа проводится во всех научно-исследовательских институтах животноводства союзных республик и во многих учебных сельскохозяйственных институтам Изучение антигенов эритроцитов осуществляется в племенных хозяйствах у всех основных видов сельскохозяйственных животных: крупного рогатого скота, лошадей, свиней, овец, птицы, пушных зверей, рыб. В последнее десятилетие в сферу иммуногенетических исследований вошло изучение антигенов белых клеток крови, спермиев и ряда других биологических объектов.
Особенности генетики эритроцитарных антигенов. Для изучения и тестирования эритроцитарных антигенов в иммуногенетике применяют методы серологических реакций: реакции гемолиза эритроцитов, агглютинации, преципитации и др. С помощью этих тестов определяют индивидуальную антигенную характеристику у отдельных особей. Эритроцитарные антигены еще называют «кровяными факторами»
Эритроцитарные антигены представляют собой сложные биополимерные макромолекулы, которые накапливаются на оболочке (строме) эритроцитов и соединяются с молекулами веществ оболочки. Структура и химический состав эритроцитарных антигенов разнообразны и характерны для каждой особи того или иного вида. Молекулы антигенов содержат мукополисахаридные комплексы, Основа биосинтеза эритроцитарных антигенов определяется действием генов и структурами ДНК и РНК.
Антигены имеют различную специфичность: видовую, групповую, типовую, патологическую, органоидную, функциональную. Антигенные особенности обусловлены последовательностью и качественными различиями аминокислот, а также особенностями строения первичной полипептидной молекулы антигена. На поверхности молекулы антигена имеются наиболее активные участки — детерминантные группы, которые определяют специфичность антигена.
Синтез каждого эритроцитарного антигена обусловлен действием одного гена. Антиген может наследоваться отдельно от других антигенов, проявляя чаще всего доминантный или кодоминантный тип наследования, и только в редких случаях наблюдается рецессивное наследование некоторых антигенов. Кроме отдельного наследования каждого антигена, наблюдается совместное наследование определенного сочетания нескольких антигенов. Это явление вызвано полным сцеплением тех генов данного локуса, которые детерминируют синтез совместно наследуемых антигенов.
Системы антигенных локусов группы крови и антигены с 1928 г. принято обозначать буквами латинского алфавита. Антигены обозначают прописными или строчными буквами латинского алфавита, но так как число антигенов велико и букв алфавита не хватает, их записывают с надстрочными или подстрочными индексами в виде штриха или цифры. Например, разные антигены получают обозначения: А, В, С, D, А1, В1 С1 D1 и т. п. Следует помнить, что А и А1 или D и D1 — это разные антигены, не связанные ни генетически, ни иммунологически друг с другом. Иногда антиген обозначают двумя буквами: строчной и прописной; так, некоторые антигены свиней записывают следующим образом: Ea, Ее, Ed.
Антигены некоторых систем образуют группы с определенной комбинацией входящих антигенов — феногруппы. Число антигенов в феногруппе системы локуса В включает 12 антигенов: В, G, К, О2, Y1, А´, В', Е3', G', К', О', Y'. Для упрощения записи таких феногрупп вводят цифровой код, поэтому указанная феногруппа обозначается буквой системы и кодовым числом: В28.
Одиночные или сцепленно наследуемые в виде постоянного сочетания антигены, которые передаются от родителей потомкам как наследственные единицы, называют группами крови. Каждая группа крови наследуется как определенная генетическая единица. В состав конкретной группы крови может входить один или несколько антигенов. Например, упомянутая феногруппа 528 из системы B крупного рогатого скота участвует в образовании группы крови со следующим антигенным составом. В, G, К, 02 ,Y1, A', E3', К'.
Контроль каждой группы крови обусловлен действием генов одного локуса и его аллелями.
Под аллелем подразумевают такой элемент генетической системы организма, который характеризует различные состояния гена определенного локуса. Ген может иметь один аллель или несколько, и тогда образуется серия аллелей данного гена. Каждое животное в генотипе соматических клеток идет два аллеля локуса — один от матери, другой от отца, В иммуногенетике аллелем служит один антиген или комплекс антигенов, передаваемый как одно целое от родителей потомку. Если отсутствует один из аллелей, то такое состояние гена в локусе считается рецессивным и тогда генотип животного выражают в виде дроби, в числителе и знаменателе которой для отсутствующего аллеля ставят прочерк. Например, при наличии одного аллельного состояния антигена А и отсутствии его второго аллеля генотип животного записывают так: А/ — или — /А.
Совокупность групп крови, контролируемых аллелями одного локуса, образует систему крови. Каждой системе крови присваивают определенное буквенное обозначение. Число уже открытых систем и входящих в каждую антигенов у животных разных видов неодинаково (табл. 41).
Системы групп крови подразделяют на простые, сложные, закрытые, открытые Если система содержит один-два антигена и имеет два аллеля — это простая система; например, у крупного рогатого скота это системы L, N.
Сложная система характерна тем, что в нее входят три антигена и более, образующих комплексные группы (системы В, С у крупного рогатого скота). Закрытые системы отличаются тем, что генотипы животных можно выявить по антигенам эритроцитов. Открытые системы — это системы групп крови» при которых генотип животного можно установить по фенотипу только у некоторых гомозигот.
Эритроцитарные антигены и группы крови изучены у ряда других видов животных: кроликов, норок, крыс, мышей, собак, уток, индеек, верблюдов, северных оленей, рыб, китов и др.
Каждая генетическая система крови определяется аллелями какого-либо одного локуса и наследуется независимо одна от другой. При этом каждый аллель определяет образование одного эритроцитарного антигена. Если локус имеет два аллельных состояния, то это вызывает формирование двух или трех генотипов и соответствующее количество фенотипов, например, си-

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
