
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Процесс дифференцировки клеток обусловлен дифференциальной экспрессией генов, когда происходит стимуляция активности одних генов и подавление (репрессия) других. Неодновременная активность различных участков молекулы ДНК была установлена при изучении структуры политенных хромосом из слюнных желез дрозофилы на разных стадиях ее развития (рис. 45). Политенные хромосомы характеризуются чередованием дисков и вздутий, называемых пуффами (от англ. puffs — вздутия). Каждому вздутию соответствует деспирализованный участок молекулы ДНК, на котором осуществляется синтез специфических молекул мРНК. Доказательством этого служит активное «поглощение» на этом участке радиоактивных «меченых» нуклеотидов. Характер и место образования пуффов меняются в различные периоды онтогенеза. Каждый диск превращается в пуфф в определенный период жизни личинки. Состояние вздутия на определенном участке политенной хромосомы обратимо, и при переходе личинки в следующую стадию пуфф превращается в диск.
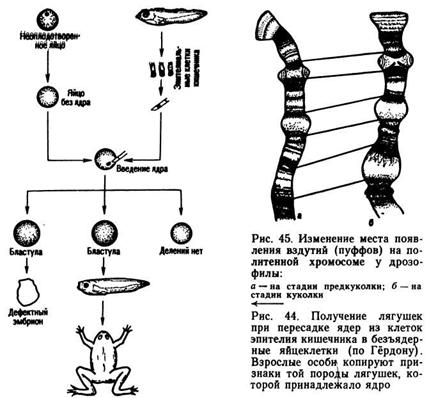
О дифференциальной экспрессии генов в онтогенезе свидетельствует также изменение состава белковых фракций на разных стадиях развития. Так, например, у человека на стадии раннего эмбрионального развития образуется гемоглобин F, состоящий из двух цепей полипептидов — α и γ. Примерно с 13-й недели развития начинает синтезироваться гемоглобин типа А, характерный для взрослого человека. Он состоит из цепей полинуклеотидов α и β. У новорожденного 70—80% составляет гемоглобин типа F, а 20 — 30% — типа А. Когда ребенок достигает возраста одного года, происходит полная замена гемоглобина F гемоглобином А. Смена соотношения типов белков в крови с возрастом наблюдается также и у животных.
В процессе дифференциации клеток и тканей происходят необратимые изменения в состоянии ДНК, хромосом, ядра и цитоплазмы, в результате чего клетка теряет тотипотентность и, хотя в ней и содержится вся генетическая информация, она не способна восстановить целый организм. В процессе дифферен-цировки и функционирования соматических клеток могут происходить морфологические изменения хромосом, значительная часть которых находится в состоянии гетерохроматина. Нарушается процесс деления клеток. Некоторые из них делятся по типу амитоза, другие — эндомитоза. В результате этого наблюдаются полиплоидизация, анеуплоидизация и другие изменения числа хромосом, образование политенных хромосом. У животных такого рода деления имеют место в клетках мальпигиевых трубок, кишечного эпителия, в клетках печени и других желез внутренней секреции.
Роль генетической информации на начальных стадиях онтогенеза. У животных в яйцеклетке до оплодотворения накапливается (в цитоплазме) большое количество рибонуклеиновых кислот всех трех типов: мРНК, рРНК и тРНК, — которые до оплодотворения находятся в неактивном состоянии. Они соединяются со специфическими белками-гистонами и образуют неактивные гранулы инфорсомы. Через несколько минут после оплодотворения часть молекул мРНК информосом освобождается от белка, поступает на рибосомы цитоплазмы яйцеклетки и начинает синтез определенных белков, необходимых %ля начального развития зиготы. Начальный период развития зиготы осуществляется под контролем генов материнского организма; мРНК яйцеклетки обеспечивает синтез белков до стадии поздней бластулы. С начала стадии гаструляции и в дальнейших процессах онтогенеза синтез белка осуществляется под контролем ядерных генов обеих родительских особей. В эмбриогенезе лягушки синтез мРНК возобновляется после 10 делений дробления, когда зародыш состоит приблизительно из тысячи клеток.
(1976) изучал первые стадии развития зародышей вьюна. Он обрабатывал оплодотворенные яйца лучами Рентгена. Дозы облучения были подобраны таким образом, чтобы прекратить деятельность ядра я нейтрализовать гены в молекулах ДНК. При облучении яиц сразу после оплодотворения или зародыша на стадии ранней бластулы развитие его шло нормально до поздней бластулы, а затем прекращалось, и наступала гибель. Следовательно, развитие эмбриона в этот период определяется иРНК, находящейся в цитоплазме клеток бластулы. Эта материнская иРНК и рРНК обеспечивают на ранней стадии дробления зиготы и бластулы синтез белков, необходимых для функционирования клеток и развития эмбриона. Развитие эмбриона прекращается на стадии гаструлы, так как для начала органогенеза нужны белки, синтез которых кодируется ядерными генами, локализованными в хромосомах материнской и отцовской особей.
При облучении эмбрионов в период гаструляции, перед началом органогенеза и даже в период поздней бластулы развитие их прекращалось сразу после облучения.
В некоторых случаях наблюдается наличие в цитоплазме яйцеклетки специальных фрагментов активной ДНК. Они синтезируют мРНК и кодируют синтез специфических белков в цитоплазме. У амфибий и рыб в цитоплазме в период созревания ооцитов и в яйцеклетках были обнаружены в большом количестве фрагменты молекул ДНК. В период созревания яйцеклеток происходит интенсивное насыщение цитоплазмы ДНК, мРНК, рРНК, тРНК, а также другими компонентами. У дрозофилы 15 фолликулярных клеток, окружающих яйцеклетку, проникают в нее цитоплазматическими выростами и насыщают цитоплазму митохондриями, ДНК, РНК, белками и другими компонентами.
Таким образом, ядро зиготы, образовавшееся в результате слияния материнского и отцовского ядер и объединения их генетической информации, в начальный период развития зародыша не оказывает влияния на дробление зиготы и образование бластулы. Эмбрион в этот период развивается за счет РНК и других компонентов, находящихся в цитоплазме яйцеклетки.
Критические периоды развития. Эмбриологи установили, что в онтогенезе, особенно на ранних стадиях развития, наблюдаются периоды, когда наиболее ярко выражена реакция эмбриона на воздействие внешних факторов. В эти периоды эмбрионы легко повреждаются, у них нарушаются процессы развития органов, что приводит к гибели эмбрионов либо к появлению уродств. Критические периоды обычно предшествуют началу соответствующего процесса органогенеза. В это время в соответствии с генетической программой развития особи усиливается синтез соответствующих белков, прекращается синтез предшествующих веществ, происходит перестройка обмена веществ в клетке. Критические периоды, как правило, наступают после поздней бластулы, когда дальнейшее развитие эмбриона осуществляется под контролем генетической информации обеих родительских особей.
Наиболее изучены внешние факторы, влияющие в критические периоды на процесс онтогенеза у рыб, птиц, амфибий, рептилий, несколько меньше — у млекопитающих.
У рыб нормальный онтогенез зависит от температуры воды и содержания в ней кислорода, причем у разных видов потребность в этих факторах различна: вьюн менее чувствителен к этим факторам, чем форель, лосось. У кур на эмбриогенез большое влияние оказывают температура и влажность воздуха в период инкубации. Эмбрионы особенно чувствительны к данным факторам на 2 — 3-й сутки инкубации, когда происходит образование системы кровообращения; на 8 — 9-е сутки — в период интенсивного морфогенеза — и на 19-е сутки, когда происходит переход зародыша к легочному типу дыхания.
Критические периоды онтогенеза определены у хомяков, морских свинок, кроликов и других животных. У крупного рогатого скота наблюдается повышение эмбриональной смертности в первые дни развития зиготы, что свидетельствует о критическом периоде.
Регуляция синтеза белков у эукариот в процессе онтогенеза. Процесс регуляции синтеза белков, разработанный Ф. Жакобом и Ж. Моно для прокариот на примере Е. coli и получивший название механизма индукции-репрессии, возможен и у высших организмов. Вместе с тем для сложноорганизованных многоклеточных эукариот характерно наличие дифференцированных органов и тканей, состоящих из узкоспециализированных клеток. В этих клетках в активном состоянии находится только та часть генетической информации, которая необходима для синтеза строго определенных белков. В дифференцированных клетках интенсивно синтезируются белки определенного состава и функций, характерных для данного органа и ткани.
Репликация ДНК. Дифференциация клеток определяет и их способность делиться по типу митоза, амитоза, эндомитоза Характер деления зависит от способности ДНК синтезировать белки, обеспечивающие репликацию ДНК и митотический цикл. В высокодифференцированных клетках, таких как нейроны, мышечные клетки, репликация ДНК не происходит довольно длительное время. Большие интервалы между делениями имеют место также в клетках печени. Вместе с тем дифференцированные клетки эпителия кишечника, костного мозга довольно интенсивно делятся и проходят полный митотический цикл.
Стабильность мРНК. В отличие от прокариот мРНК у эукариот, особенно в клетках животных, относительно стабильна и может длительное время служить матрицей белкового синтеза, а также сохраняться в цитоплазме в виде информосом. Так, например, у человека длительность жизни ретикулоцитов до превращения их в эритроциты равна шести суткам. Ядра у них отсутствуют, но синтез специализированная молекул белка в них протекает на мРНК, образовавшихся в ядрах на предшествующей стадии нормобласта. Таким образом, у высших организмов возможно образование безъядерных клеток, которые могут нормально функционировать за счет ранее синтезированных мРНК. Аналогично осуществляется синтез белка на ранее синтезированной иРНК и в клетках, содержащих неактивные ядра, как, например, в эритроцитах птиц, спермиях и других дифференцированных клетках. Таким образом, для аукариот характерно иногда довольно продолжительное неодновременное протекание процессов транскрипции и трансляции.
Каскадная регуляция активности генов. Она заключается в том, что в клетке происходит одновременное включение или выключение большой группы генов, локализованных в разных молекулах ДНК, разных хромосомах. Эта регуляция осуществляется под воздействием специализированных весьма разнообразных сигнальных веществ, активно синтезируемых в клетках других тканей и поступающих в клетки данной ткани.
Гормональная регуляция. У высших животных важное значение имеет гормональная регуляция активности генов. Гормоны вырабатываются железами внутренней секреций и активируют синтез соответствующих белков. Они могут иметь белковую и небелковую природу, но синтез каждого из них осуществляется под генетическим контролем ДНК. Выделяясь из соответствующих желез в кровь, гормоны разносятся по всему организму, вступают в контакт с соответствующими клетками и активируют их гены. Гормоны контролируют многие процессы онтогенеза: рост, органогенез, морфогенез, метаморфоза у насекомых и амфибий, наступление половой зрелости и другие процессы.
Оптимальное количество гормонов является непременным условием нормального развития и существования организма, Недостаток одного или нескольких гормонов приводит к нарушению процесса развития, иногда к стерильности особи. Избыток гормонов нарушает процесс обмена веществ в организме, обусловливает эндокринные расстройства.
Ряд гормонов влияет непосредственно на ДНК дифференцированных клеток и регулирует синтез специфических белков, Гормоны являются либо индукторами, либо супрессорами синтеза мРНК, или изменяют проницаемость клеточной мембраны для специфических индукторов синтеза мРНК.
Гормоны могут соединяться с молекулами ферментов и изменять их активность. Об активации ферментами геной, влияющих на процессы метаморфоза, свидетельствуют следующие эксперименты. У двукрылых насекомых был выделен специальный гормон развития. Введение его личинкам способствовало быстрому их превращению в куколки. При добавлении в питательную среду гормона щитовидной железы головастики быстро превращались в лягушек, аксолотль — в амблистому.
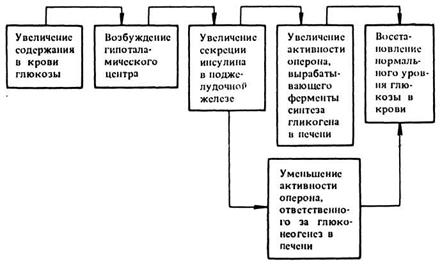
Примером регуляторной деятельности гормона может служить инсулин — наиболее изученный гормон поджелудочной железы. Инсулин — белок, состоящий из одной полипептидной цепи, содержащей 51 аминокислоту. Благодаря инсулину в крови поддерживается необходимая концентрация глюкозы, имеющей огромное значение в жизнедеятельности и развитии организма. Инсулин регулирует работу генетического аппарата клеток печени, в которых синтезируются ферменты, необходимые для нормального течения двух противоположных процессов — синтеза глюкозы из неуглеводистых веществ и гликолиза глюкозы и синтеза из нее гликогена. Оптимальная концентрация глюкозы в крови поддерживается соотношением комплекса ферментов двух этих систем.
Инсулин активирует оперон, содержащий три структурных гена, синтезирующих ферменты, необходимые для гликолиза и синтеза гликогена. В то же время инсулин является репрессо-ром четырех генов другого оперона, влияющего на синтез глюкозы. приводит следующую схему регуляции активности генов печени при поступлении с пищей в организм человека большого количества сахара:
Продуктивность различных пород животных зависит от активности различных гормонов. У мясных пород свиней гормон соматостатин активирует белковый синтез, необходимый для более эффективного использования кормов и увеличения живой массы. Особенно большое влияние на регуляцию работы генетического аппарата клетки оказывает гормон роста, вырабатываемый гипофизом. Этот гормон является индуктором синтеза белков во многих клетках одновременно. Например при удалении гипофиза резко уменьшается синтез белков, необходимых для нормального роста.
Например, у белых крыс при гипофизэктомии вдвое уменьшается синтез белков и количество полисом по сравнению с контролем. При введении этим крысам гормона роста синтез мРНК, рРНК и белков возвращается к норме.
Наряду с хорошо известными гормонами в организме существуют и другие высокоспециализйрованные индукторы.
Большое влияние на активность соответствующих генов оказывают цитоплазма дифференцированных клеток, а также белки-гистоны, В процессе дифференциации клетка приобретает способность реагировать только на определенные раздражители, в результате чего она синтезирует только те белки, которые необходимы для ее дальнейшего функционирования, жизнедеятельности и дифференцировки. Характерно, что свойство генетического аппарата дифференцированной клетки синтезировать специфические белки клетка сохраняет и в последующих клеточных поколениях. Таким образом, увеличение размеров ткани за счет деления клеток не изменяет работу генетического аппарата, и дифференцированные клетки синтезируют те же специфические для данной ткани белки.
Генотип и фенотип. Ген как дискретная единица наследственности реализуется в процессе синтеза ферментов или структурных белков. На молекуле ДНК транскрибируется мРНК, на которой в цитоплазме образуются полипептидные цепи. В посттрансляционный период полипептиды претерпевают различные перестройки: соединяются друг с другом, с молекулами небелковой природы, приобретают вторичную, третичную или четвертичную структуру. Они участвуют в развитии и формировании органов и признаков, выполняя специфические функции ферментов, структурных или транспортных белков. В свою очередь, каждый этап реализации наследственной информации в процессе биосинтеза контролируется сложной ферментативной и регуляторной системой. Результаты проводимых исследований свидетельствуют о дискретном характере наследственности, когда характер проявления одного признака контролируется одним, двумя или большим числом генов, локализованных в определенных участках молекулы ДНК и хромосомы.
Вместе с тем генотип нельзя рассматривать как мозаику дискретных единиц — генов. В процессе онтогенеза он проявляет себя как единая система, регулирующая все процессы развития органов и признаков.
Фенотип особи определяется всей суммой индивидуальных Признаков, доступных наблюдению или анализу. Он также имеет дискретную природу, так как каждому организму свойственны специфические для него признаки. С другой стороны, фенотип представляет собой единое целое и нарушение строения одного органа сказывается на жизнеспособности всего организма. Фенотип особи складывается в онтогенезе под контролем генотипа и под влиянием условий среды. В эмбриональный период онтогенеза органогенез осуществляется на основе тесной взаимосвязи процесса заложения и развития органов
Пенетрантность и экспрессивность генов. Проявление действия гена может иметь различный характер, и фенотипическое проявление его может варьировать по степени выраженности признака. Один и тот же признак может проявляться или не проявляться у особей родственных групп. Это явление называется пенетрантносяью гена, Пенетрантность определяют по проценту особей в популяции, у которых данный ген проявился, Если он проявится у всех обследованных животных или растений, то пенетрантность будет составлять 100%, если у части особей, то определяют их процент — 80, 75 % и т. д.
Экспрессивность гена характеризует фенотипическое проявление гена по реакции сходных генотипов на конкретные условия внешней среды. Действие одних генов в онтогенезе может быть более или менее константным, стойким в своем проявлений или варьировать в зависимости от внешних условий. Рецессивные гены, которые в обычных условиях в гетерозиготном состоянии фенотипически не проявляются, могут проявиться при измененных условиях.
Примером экспрессивности и различного фенотипического проявления гена может служить окраска меха у кролика, определяемая серией множественных аллелей гена С.
Гималайская окраска меха определяется аллелем ch и фенотипически проявляется в, белой окраске меха, но с черной окраской кончиков лап, ушей, носа и хвоста (рис. 46).

Окраска меха зависит от биохимических реакций, протекающих в клетках кожи, контролирующих выработку меланина, и от температуры окружающей среды. Кролик, выращенный при температуре выше 30 °С, оказывается сплошь белым. Если выщипать небольшой участок белой шерсти и систематически его охлаждать, то на нем вырастет черная шерсть. Пониженная температура влияет на активность генов, контролирующих выработку определенных ферментов,
Важное значение для характера проявления генов имеют наличие и активность генов-модификаторов, определяющих степень экспрессивности генов в зависимости от условий среды. В постэмбриональный период, когда идет процесс формирования конкретных признаков, присущих данной особи, генотип проявляется в фенотипе как система взаимосвязанных генов. В этой системе развитие одного признака может зависеть от взаимодействия многих генов и один ген может влиять на развитие и проявление нескольких признаков.
Плейотропия. Явление одновременного влияния одного наследственного фактора — гена — на несколько признаков называют плейотропией. Плейотропное действие гена может быть как положительным, так и отрицательным. Практически всегда при анализе связей гена и признака можно обнаружить явление плейотропии. Особенно четко плейотропия проявляется при изучении фенотипических изменений признаков, вызываемых мутацией одного гена. У животных и человека мутация одного гена может обусловить целый комплекс патологических изменений фенотипа, называемых в медицине синдромами.
В медицине и ветеринарии наиболее изученными являются так называемые синдромные и биохимические плейотропии. При синдромной плейотропии один мутантный ген o6yсловливает комплекс повреждений в процессе раннего эмбрионального развития. Примером синдромной плейотропии может служить рецессивная мутация гена, кодирующего фермент галактозо-1-фосфатуридилтрансферазу, необходимый ребенку для усвоения молочного сахара. Эта мутация обусловливает заболевание галактоземией и оказывает плейотропный эффект на комплекс признаков. У больного проявляются такие пороки развития, как слабоумие, слепота, цирроз печени. Предупредить развитие заболевания можно, если больного перевести на искусственную диету, не содержащую молочного сахара. На первый взгляд эти дефекты не связаны между собой, хотя обусловлены мутацией одного гена.
Примером биохимической плейотропии, послужившей моделью для изучения сложных взаимодействий структурных генов и характера их проявления в фенотипе особи, могут служить мутации генов, контролирующих метаболизм аминокислот фенилаланина и тирозина. Нарушение этого метаболизма обусловливает патологические изменения целой серии признаков у человека и такие заболевания, как фенилкетонурия, алкаптонурия, кретинизм, альбинизм (рис. 47).

Фенилкетонурия — тяжелое наследственное заболевание человека. Впервые оно описано в 1934 г., но только через 19 лет, в 1953 г., было установлено, что фенилкетонурия является следствием мутации гена, ответственного за синтез фермента, обеспечивающего гидролиз аминокислоты фенилаланина и синтез тирозина. Неспособность генотипа вырабатывать этот фермент приводит к тому, что поступающая с пищей аминокислота фенилаланин накапливается в плазме крови, а затем — в мозге. Избыток ее определяет плейотропный эффект, затрагивающий патологическое развитие комплекса признаков, и у больных детей развивается умственная отсталость, а затем и неполноценность, потеря речи, отсутствие координации движений.
После того как были установлены причины фенилкетонурии, разработаны методы диагностики у новорожденных и с помощью специальной диеты получена возможность предотвратить это заболевание.
Алкаптонурия проявляется в возрасте 40 лет и старше и характеризуется патологическими изменениями суставов конечностей, позвоночника, потемнением мочи. Причиной комплекса этих патологий является то, что у больных в печени не вырабатывается фермент оксидаза, необходимый для метаболизма гомогентизиновой кислоты. В организме происходит ее накопление, и развивается тяжелое заболевание.
Тирозиноз — заболевание, обусловленное нарушениями в метаболизме
аминокислоты тирозина. Накопление в организме избытка данной аминокислоты и ее метаболитов обусловливает задержку в развитии младенца,
кретинизм, слабоумие, патологию почек и печени.
Причиной альбинизма является не избыток, как в предыдущих случаях, а недостаток продукта - меланина, поэтому патология бывает выражена в раннем возрасте, уже у новорожденных. Причиной альбинизма бывает потеря организмом способности синтезировать меланин из тирозина из-за отсутствия фермента тирозиназы. Этот фермент в норме катализирует образование пигмента меланина в клетках кожи, волос, радужной оболочки глаз, Отсутствие его блокирует образование меланина, и люди рождаются альбиносами.
Коррелятивные связи органов. В формировании фенотипа важное значение имеют коррелятивные связи органов и признаков. (1884—1963) выделял следующие типы коррелятивных связей: геномные, морфологические и функциональные. Геномные связи обусловлены действием генотипа как целостной системы в процессе онтогенеза. Морфологические корреляции наследственно обусловлены и возникают в результате передачи веществ или возбуждений от одной части организма к другой. Функциональные корреляции являются следствием функциональной связи между органами. считал, что в онтогенезе особую роль играют морфогенетические корреляции, благодаря которым развитие организма и формирование фенотипа осуществляются как единый процесс. Выявление коррелятивных связей и их роли в развитии признаков имеет важное значение в генетике и селекции животных.
Генетические основы воспроизведения и долголетия животных. Воспроизведение потомства и долголетие обусловлены сложными генетическими процессами, осуществляемыми на фоне взаимодействия организмов с разнообразными факторами среды.
Под воспроизведением животных понимают способность организмов давать полноценный приплод, определяемый качеством родительского поколения. Воспроизведение и продуктивность являются тесно связанными между собой биологическими процессами, на уровень которых влияет состояние животных, их наследственность и условия жизни.
Интенсивность воспроизведения, как и показатели продуктивности, входит в комплекс селекционных признаков. В основе селекционного эффекта, означающего повышение интенсивности воспроизведения, лежат условия жизни и наследственные особенности размножающихся особей и их потомков. Опираясь на эти факторы, можно методом селекции достигать повышения воспроизводительности животных.
Практическими показателями нормального процесса воспроизведения животных в ряде поколений могут служить: биологическое и хозяйственное долголетие животных, оплодотворяемость самок и спермояродукция самцов, пренатальная и постэмбриональная жизнеспособность, сопровождающаяся приспособленностью потомства к условиям внешней среды и элементам технологии, способность длительное время проявлять высокие продуктивные качества при интенсивном воспроизведении. Однако даже при создании необходимых условий жизни могут быть срывы и снижение воспроизводительной функции, проявляться повышенная пренатальная смертность, снижение жизнеспособности и продуктивности в последующие периоды онтогенеза.
Основу функции воспроизведения определяют процессы, обусловленные наследственностью животного, особенностями его генетического аппарата и биосинтеза белковых молекул, в частности таких, как антитела, иммуноглобулины, ферменты, антигены, гормоны и другие активные биологические соединения, то есть комплекс иммунологических факторов.
Иммунологическая обусловленность уровня воспроизведения. В последние годы установлено, что в процессах гаметогенеза, оплодотворения и взаимоотношений между эмбрионом и материнским организмом важную роль играют взаимосвязи иммунной системы самца, самки и эмбриона. В основе взаимосвязей — соотношение антигенов половых клеток родителей и антител материнского организма и зародыша.
Антигенными свойствами обладают клетки, ткани и жидкости тела, в частности клетки семенников и яичников, секреты добавочных половых желез самцов, клетки молочной железы и половой системы самок, органоиды половых клеток (акросома) ядро, прозрачная оболочка и др.). Ответная реакция на антигены выражается синтезом антител, то есть иммунным ответом.
По инициативе болгарских ученых в 1967 г. была проведена. Первая международная конференция по иммунологии воспроизведения, в которой участвовало 40 стран мира, В 1980 г. в Париже проведен первый международный конгресс по иммунологии воспроизведения. В СССР организован Советский комитет по иммунологии воспроизведения и иммуногенетике и в 1977 г. проведен первый симпозиум, Исследования по иммунологии воспроизведения в нашей стране ведут академик и профессор .
Иммунные процессы, влияющие на воспроизведение, проявляются на различных уровнях и этапах этой функций. В комплекс процессов входят следующие компоненты: а) формирование иммунных особенностей гамет, образующихся в процессе спермато - и оогенеза; б) взаимодействие иммунных систем половых продуктов самца и самки после осеменения и при продвижении спермы в половых путях; в) иммунные процессы, происходящие в период собственно оплодотворения и образования зиготы; г) иммунные взаимоотношения организма матери и плода в пренатальный период. Каждый из этих компонентов иммунного комплекса имеет свою специфику, иммунную особенность и силу влияния на качество приплода.
Для выявления иммунных взаимосвязей антиген — антитело используют методы реакции преципитации, агглютинации, иммунофореза и др. Важное значение в системе биотехнологии приобретают пересадки зигот от самок-доноров к самкам реципиентам, а также при использовании искусственного осеменения в условиях in vitro и in vivo.
Иммунные процессы при гаметогенезе. В процессе сперматогенеза у спермиев формируется иммунная система, включающая до 18 антигенов, что обусловлено их сложной химической структурой. Дополнительным источником антигенности спермия служат жидкости добавочных половых желез.
Антигены головки и хвоста спермия различны. Выявлено общее сходство некоторых антигенов между спермиями быка, барана и хряка. Иногда выявляется сходство некоторых антигенов семенной жидкости с антигенами крови самца.
Антигенная структура спермия формируется постепенно в процессе сперматогенеза. Так, генетическая обусловленность его антигенности зависит от хромосомного аппарата гаплоидных сперматид. Кроме этого, антигенность гамет создается еще и за счет образования липопротеидной оболочки спермия из секретов придатков семенников.
В акросоме спермия иммуноантигенность проявляют такие ферменты, как гиалуронидаза, дегидрогеназа и другие, оказывающие влияние на оплодотворяющую способность спермия. Так, гиалуронидаза способствует освобождению яйцеклетки от клеток яйценосного бугорка и лучистого венца, что помогает спермию достигнуть прозрачной оболочки ооцита, Лактатдегидрогеназа влияет на дыхательный процесс спермия и его подвижность.
На поверхности спермиев выявлены антигены, обусловленные функцией генов, вызывающих гистонесовместимосты Их действие может приводить к отторжению пересаженных участков ткани или органов. В современной зоотехнии применяемая пересадка зигот или эмбрионов в качестве приема генетической инженерии также может сопровождаться реакцией гистонесовместимости между пересаженной зиготой или эмбрионом от самки-донора к самке-реципиенту. В результате происходит гибель пересаженных элементов. Следовательно, иммунное состояние самки-реципиента влияет на успех пересадки.
У мышей выявлен антиген гистосовместимости, который определяется геном, расположенным на Y-хромосоме Опыты и показали, что в результате обработки крольчих спермой кролика, подвергнутого воздействию антисывороткой, иммунной к клеткам кожи самца, наблюдается существенный сдвиг в соотношении полов у приплода. Это позволило авторам высказать гипотезу о возможности использования антигенных свойств спермы для регулирования соотношения полов в потомстве
Для нормального воспроизведения важно, чтобы в организме самца не было процесса образования антител в отношении собственных спермиев, то есть, чтобы отсутствовала аутоиммунность с образованием аутоантител, наличие которых приводит к агглютинации (склеиванию) головок собственных спермиев и тем самым к утрате их оплодотворяющей способности. Образование аутоантител у самца может вызываться различными внешними факторами, например перегревом или переохлаждением семенников, их ушибами и другими травмами органа. Учитывая отрицательное влияние аутоиммунных антител, необходимо тщательно оберегать самцов от травм.
Аутоантитела также могут синтезироваться клетками, в которых произошла мутация гена, вызывающая искажение реакции на антигены собственного организма. Аутоиммунизация сопровождается повреждением акросомы головки спермия, она набухает и даже утрачивается. Патологические спермин часто присутствуют в придатках семенников и эякуляте.
При аутоиммунизации количество патологических гамет в придатках самцов достигало 49% и в эякуляте 46, а у нормальных самцов около 30% (Боярский, 1978).
Появление аутоиммунности у здоровых самцов под влиянием внешних факторов, вызывая патологию в состоянии акросомы спермия, приводит к снижению оплодотворяемости самок. Для диагностики аутоиммунности в практике животноводства можно использовать степень реакции оседания спермиев (РОС), появляющейся в антисыворотке данного самца.
В опытах и др. (1960) из 59 обследованных быков-производителей реакция оседания была выявлена у шести быков Высокий процент стельности у коров отмечен при использовании быков с отрицательной реакцией оседания спермиев На практике важно осуществлять проверку производителей для систематической оценки спермопродукции.
предложен метод определения биологической (оплодотворяющей) способности спермы по состоянию акросомы спермия. Для этого просматривают капли спермы под микроскопом с темнопольным конденсором. У неполноценных спермиев акросома не светится или даже отсутствует, а у нормальных передний край акросомы ярко освещен. Инструкция по оценке состояния акросом уже внедряется в практику скотоводства и позволяет оценивать быков-производителей по состоянию их спермопродукции в динамике в период их использования.
Присутствие многообразных антигенов у спермиев и в семенной жидкости придатков может служить фактором, снижающим эффект воспроизведения в результате выработки половой системой самки антител к веществам антигенного типа в продуктах воспроизводительной системы самца, а также и уже развивающемуся зародышу. В половом тракте самца существуют защитные механизмы, предупреждающие проявление антигенного влияния половых продуктов самца и его аутоиммунность. Например, защитными свойствами обладают липопротеидный покров спермиев и слабокислая реакция эпидидимиса. Кроме этого, капилляры извитых канальцев семенника непроницаемы для антител, что предотвращает возможность иммунной реакции антиген — антитело на уровне сперматогенеза. Этот процесс защиты в семеннике называется гематотестику-лярным барьером.
Антигенные свойства яичника и половой системы самки. Закладка фолликулярного слоя с оогониями у самок происходит в эмбриональном периоде, поэтому антигенные свойства половых клеток самок проявляются уже в пренатальном периоде. Антигены были выявлены в яичнике, яйцеводах, матке, желтых телах и фолликулярной жидкости. В яичнике обнаружено около 10 антигенов, которые отсутствуют в других тканях тела. Такое число антигенов яичника могло бы вызвать реакцию аутоиммунных процессов, но охрана антигенной системы зрелого ооцита от антител, которые может синтезировать организм самки, достигается за счет наличия сосудистой оболочки фолликула и большим числом слоев фолликулярных клеток, не пропускающих прорастания капилляров, несущих клетки, синтезирующие антитела. Аналогичную защитную роль от аутоантител выполняют фолликулярная жидкость и прозрачная зона ооцита, Прозрачная зона ооцита содержит вещества, которые служат имунной защитой в период прохождения ооцита, зиготы или бластоцисты по половой системе самки. Она является важным элементом, обеспечивающим сохранение яйца, зиготы, бластоциста от возможного иммунного воздействия антител, выделяемых иммунной системой оплодотворенной самки, и тем самым повышает эффект воспроизведения.
После введения семенной жидкости самца в половые пути самки наступает неспецифическая иммунная реакция ее организма в виде фагоцитоза. При этом сегментоядерные нейтрофильные лейкоциты устраняют мертвых и слабых спермиев.
Реакция организма самки на введенную семенную жидкость самца сопровождается появлением в крови естественных антител типа агглютининов, лизинов. Если самка несколько раз была осеменена и беременность не наступила, то концентрация агглютининов в ее крови увеличивается и может превышать критический уровень антител (1:15, 1:32). Повышение титра агглютининов многократным осеменением может достигать соотношения 1:639. Такое соотношение антител и антигенов спермиев приводит к снижению оплодотворяемости самок.
Повышение титра агглютинации может быть вызвано недостатком витаминов A, D, E и микроэлементов (кобальта). Оно наблюдается в первый месяц после отела, поэтому осеменение в первую охоту, то есть менее чем через 30 дней, часто сопровождается прохолостом коровы. Естественные антитела к спермиям выявлены у самок не только в крови, но и в слизи половых путей. Эти антитела в основном относят к группе иммуйоглобулинов, в частности к IgG.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
