
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Контрольные вопросы. 1. Изменение функции генов в онтогенезе животных: экспрессия, пенетрантность, 2. Критические периоды развития у животных разных видов. 3. Регуляция синтеза белков в процессе онтогенеза. 4. Гормональная регуляция и генетические процессы онтогенеза. 5. Антигенные структуры в гаметах и половой системе самок и самцов. 6. Иммунные процессы, происходящие при оплодотворении. 7. Иммунные процессы взаимоотношений матери и плода. 8. Биотехнология интенсификации воспроизведения у животных, современные методы.
ГЛАВА 10. МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
Живым организмам, независимо от их генетической организаций, наряду с наследственностью свойственна изменчивость. Наследственным изменениям свойств и признаков у микроорганизмов, растений, животных и человека всегда уделялось большое внимание. Ч. Дарвин придавал им большое значение в эволюции и селекции. Одним из широко известных примеров наследственных изменений, описанных Дарвином, является рождение в 1791 г. на ферме Анкон в штате Массачусетс в США коротконогой овцы, родоначальницы анконской породы (рис.48). Он описывает также случаи появления однокопытных животных у свиней и многие другие.

В 1899 г. вышла книга русского ботаника, в то время профессора Томского университета, (1861— 1900) «Гетерогенез и эволюция», в которой приведен ряд примеров наследственной изменчивости признаков у растений как источника происхождения видов. Термин «мутация» был введен в генетику Г. де Фризом, голландским ученым, который в течение многих лет (1886—1901) изучал явление наследственной изменчивости у растения энотеры (Oenothera lamarkiana). После тщательного обобщения своих наблюдений он разработал мутационную теорию, которую сформулировал в книге «Мутации и периоды мутаций при происхождении видов» (1901) 'Мутациями (от лат. mutatio — изменение, перемена) называют наследственные изменения признака, органа или овойства, обусловленные изменениями наследственных структур. Процесс возникновения мутаций называется мутагенезом. Мутагенез может быть спонтанным, когда мутации возникают в природе без вмешательства человека, и индуцированным, когда мутации вызывают искусственно, воздействуя на организм специальными факторами, называемыми мутагенами. Растение, животное, микроорганизм, у которых произошла мутация, называют мутантами.
Мутации — закономерное генетическое явление, характеризуемое следующими особенностями. 1) мутационные изменения обусловлены изменением наследственных структур в половых или соматических клетках и могут воспроизводиться в поколениях, то есть являются наследственными; 2) мутации возникают внезапно у единичных особей, носят случайный, ненаправленный характер, могут быть рецессивными и доминантными; 3) мутации могут идти в разных направлениях, затрагивать один или несколько признаков и свойств, могут быть ценными, полезными или вредными. Как, указывают Р. Ригер и А. Михаэлис (1967), мутации, снижающие выживаемость мутантов более чем на 10%, вредны для природных популяций. В сельскохозяйственной практике ценность мутации определяется ее значением для селекции; 4) одни и те же мутации могут возникать повторно.
В зависимости от того, изменением каких наследственных структур обусловлена мутация, принята следующая их классификация:
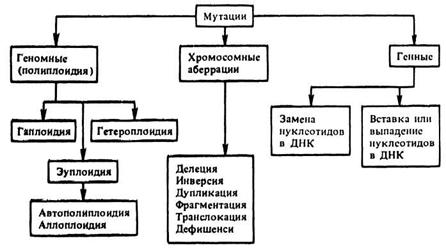
Полиплоидия. В широком смысле этого слова полиплоидией называют геномную мутацию, обусловленную изменением числа хромосом в клетках, а также процесс возникновения или создания геномных мутантов (полиплоидов).
Полиплоидные формы в природе могут возникать по ряду причин, например из-за нарушения митоза, в результате которого происходит неравное расхождение хромосомом в анафазе, отсутствие цитокинеза, нарушение функций митотического аппарата; это может наблюдаться и в результате образования и слияния при оплодотворении нередуцированных гамет, образовавшихся при нарушении мейоза, а также митотического деления зиготы или соматических клеток в начальные периоды эмбриогенеза. В зависимости от того, в каких клетках происходит изменение числа хромосом, различают соматическую, мейотическую или зиготическую полиплоидию. В природе полиплоиды чаще всего возникают либо в результате слияния нередуцированных гамет (мейотическая полиплоидия), либо в результате нарушения первого деления эиготы (зиготическая полиплоидия). Индуцированные полиплоиды чаще всего получают, воздействуя на митотическое деление клеток меристемы (органообразовате-льной ткани у растений), точек роста (соматическая полиплоидия).
С этой целью точки роста прорастающих семян или веге-тирующих растений в течение 1—5 ч обрабатывают слабым раствором колхицина (0,01—0,25%). Алкалоид колхицин блокирует развитие митотического аппарата клетки, поэтому в ней не происходит расхождения сестринских хромосом к полюсам и цитокинеза и число хромосом в клетке удваивается. Процесс удвоения числа хромосом в клетке может продолжаться до тех пор, пока будет действовать раствор колхицина.
Полиплоидия — явление, широко распространенное в природе, особенно среди растительных организмов. Многие дикорастущие и культурные виды растений являются спонтанными полиплоидами. В зависимости от климатических условий произрастания число полиплоидных видов растений может колебаться от 35 до 85%. Многие виды покрытосеменных растений образуют полиплоидные ряды в пределах одного рода. Полиплоидным рядом называют виды одного рода, у которых число хромосом увеличивается кратно гаплоидному. Например, полиплоидный ряд пшеницы (род Triticum) содержит серию видов, четко различимых по числу хромосом:
диплоидные виды (2n=14) —Т. urartu, T. monococcum и др.;
тетраплоидные виды (2n=28) —Т. dicoccum, T. durum, T. turgidum, T. polonicum и др.;
гексаплоидные виды (2n=42) — Т. aestivum, Т. tompactum, Т. spelta, Т. persicum, T. vavilovii и др.
Полиплоидный ряд картофеля (род Solatium) включает виды, содержащие в клетках 24, 48, 72, 96, 120 и 144 хромосомы, полиплоидный ряд щавеля (род Rumex)—20, 40, 60, 80, 100, 120 хромосом.
Наименьшее гаплоидное число хромосом каждого полиплоидного ряда называется его основным числом и обозначается буквой л. Например, у пшеницы основное число хромосом полиплоидного ряда X=7, у картофеля Х=12, у щавеля X=10. Совокупность хромосом основного числа полиплоидного ряда называется геномом. В зависимости от плоидности каждый вид содержит один или несколько геномов Так, растения мягкой пшеницы Г. aestivum имеют гаплоидный набор хромосом n = 21, то есть содержат 3 генома, обозначаемых буквами Л, В и D.
Гаплоидия — геномная мутация, в результате которой возникают гаплоиды — организмы с редуцированным (одинарным) числом хромосом. В клетках гаплоидов содержится только половина соматического набора хромосом (я), присущего данному виду, то есть такое же число хромосом, как и в нормальных половых клетках — гаметах. Гаплоиды могут возникать спонтанно и могут быть получены индуцированно Гаплоиды бесплодны, но могут размножаться партеногенетически и сохраняться при вегетативном размножении. Гаплоидные мутации используют в селекции высших растений. Если у гаплоида удвоить число хромосом с помощью раствора колхицина или другим методом, то можно получить гомозиготное по всем генам, нормально плодовитое диплоидное растение.
Эуплоидия (истинная полиплоидия) — геномная мутация, в результате которой возникают эуплоиды — организмы, в клетках которых содержится более двух гаплоидных наборов хромосом одного вида или происходит соединение и кратное увеличение хромосомных наборов разных видов. Различают автополиплоидию и аллополиплоидию
Автополиплоидия — процесс возникновения автополиплоидов — организмов, в клетках которых содержится более двух гаплоидных наборов хромосом, присущих данному виду. В зависимости от числа хромосомных гаплоидных наборов различают триплоиды, в клетках которых содержится 3n число хромосом, тетраплоиды (4n), пентаплоиды (5n), гексаплоиды (6n) и т. д.
Впервые явление кратного увеличения числа хромосом в клетках было описано профессором МГУ И. И Герасимовым, наблюдавшим полиплоидизацию у водоросли спирогиры в 1890 г. В 1916 г. это явление наблюдал Г. Винклер и дал ему название «полиплоидия».
Автополиплоидия обусловливает изменение морфологических признаков и свойств, присущих исходным диплоидным растениям. У полиплоидов в первую очередь увеличиваются размеры ядра и клетки в целом, а также количество органоидов цитоплазмы — пластид, митохондрий, рибосом. Для каждого вида растений существует определенный оптимальный уровень плоидности, то есть такое кратное гаплоидному число хромосом, при котором растения имеют наиболее высокую жизнеспособность и продуктивность. Так, для сахарной свеклы и арбуза оптимальным является триплоидный уровень (у арбуза Зn=33, у свеклы 3n = 27), для ржи, гречихи, редиса, турнепса — тетраплоидный. У этих растений при оптимальном уровне плоидности увеличиваются размеры листовых пластинок, длина и толщина стебля. Чашелистики, лепестки венчика, пыльцевые зерна, плоды и семена у автополиплоидов крупнее, чем у исходных диплоидных растений. Полиплоидные сорта ряда культурных растений получили широкое распространение: тетраплоидная рожь сорт Белта и Ленинградская тетра; гречиха — Искра, Эмка (Польша), Пенкрад (Канада); редис — Новосибирский тетра; клевер — Тетраплоидный ВИК (СССР), Ульва 090, Вейбуллский (Швеция), Трипо (Норвегия); триплоидные гибриды сахарной свеклы — Кубанский полигибрид 9, а также Белоцерковские гибриды 1 и 2 и др.
Особенности мейоза и наследования признаков у автополиплоидов. У диплоидного организма в профазе I мейоза происходит нормальная конъюгация двух гомологичных хромосом и образование бивалентов. У тетраплоидных организмов в клетках содержится четыре гомологичных хромосомы, что приводит в профазе I мейоза к нарушениям процесса конъюгации и образованию наряду с бивалентами увивалентов, тривалентов и тетравалентов. Унивалентами называют одиночные хромосомы в пахитене профазы I, тривалентами—ассоциации трех, тетравалентами — четырех гомологичных хромосом. В результате такого нарушения процесса конъюгации хромосом образуются гаметы и зиготы с числом хромосом, некратным гаплоидному. Так, например, тетраплоидные растения сахарной свеклы, в клетках которых содержится 4n = 36 хромосом, могут образовать гаметы, содержащие от 13 до 23 хромосом, при слиянии которых в процессе оплодотворения образуются зиготы, содержащие от 26 до 46 хромосом. Зиготы, содержащие несбалансированное число хромосом, как правило, не реализуются в семена и плоды, поэтому одним из существенных недостатков искусственно получаемых автополиплоидов являются пониженная семенная продуктивность и череззерница. При длительном размножении и тщательном отборе нормально оерненных растений в процессе селекции достигаются относительная сбалансированность хромосомного набора и стабильная семенная продуктивность у автополиплоидных растений.
При нормальной конъюгация хромосом и образовании бивалентов наследование признаков у полиплоидных растений осуществляется значительно сложнее, чем у диплоидных. В качестве примера можно привести наследование высоты растений у диплондов и тетраплоидов гречихи. У гречихи ген D обусловливает высокорослый тип растений с неограниченным ростом Рецессивный аллель этого гена контролирует так называемые детерминантные (карликовые) формы, у которых на вершине стебля вместо точки роста образуется обычная цветочная кисть. При скрещивании диплоидных гомозиготных растений DDXdd в F1 все растения имеют неограниченный тип роста, бывают высокорослыми, а в F2 наблюдается расщепление в отношении 3: 1.
При скрещивании тетраплоидных растений» имеющих генотипы DDDDxdddd, при отсутствии нарушений в мейозе все растения F1 будут высокорослыми (DDad). Они образуют 3 типа гамет в следующем соотношении: 1DD : 4Dd: 1dd. При равновероятном слиянии этих гамет в процессе оплодотворения у тетраплоидных растений могут в F2 образоваться следующие генотипы: 1DDDD : 8DDDd : 18DDdd : 8Dddd : 1ddd. При полном доминировании в F2 может наблюдаться расщепление по фенотипу в соответствии 35 высокорослых: 1 карликовый. Еще более сложно наследуются у тетраплоидов признаки, обусловленные взаимодействием неаллельных генов по типу комплементарности и полимерии.
Аллополиплоидами называют растения, в кариотипе которых содержатся удвоенные наборы хромосом разных видов и родов Они могут возникать в природе или могут быть получены искусственным путем при удвоении числа хромосом у межвидовых или межродовых гибридов. Аллогюлиплоиды, созданные в результате удвоения числа хромосом у растений, полученных от скрещивания особей, относящихся к двум разным видам или родам, называются амфидиплоидами (от греч. amphi — оба). Если аллополиплоид содержит удвоенные числа хромосом трех видов или родов, его называют аллртриплоидом.
Аллополиплоидам обычно присущи признаки и свойства исходных диплоидных родительских форм в различных сочетаниях, как это обычно бывает при межвидовой и межродовой гибридизации. Полиплоидизация позволяет восстановить плодовитость, так как межвидовые и особенно межродовые гибриды, как правило, бесплодны.
Первые амфидиплоиды были получены Г Д. Карпеченко в 1924 г. Он скрещивал редьку Raphanus sativus (2n=18) с капустой Brassika oleracea (2n =18). Хотя эти виды содержат одинаковое число хромосом, но хромосомы не конъюгируют и растения F1 были бесплодными, хотя и имели мощное развитие. Но единичные гаметы содержали нередуцированное число хромосом (9B+9R). При слиянии таких гамет растения F2 содержали 36 хромосом (18B+18R). У них нормально протекал мейоз, они были плодовитыми, мощно развитыми и давали константное потомство. Растения сочетали признаки обоих видов. редечно-капустные амфидиплоиды предложил называть «Рафанобрассика» (Raphanobrassika).
Важное практическое значение имеют амфидиплоиды, полученные путем удвоения числа хромосом у пшенично-ржаных межродовых гибридов, названных, по предложению рева, Тритикале. Они могут быть окташюидами (2n = 56) и гексаплоидами (2n = 42). Тритикале является ценной зернокормовой культурой. Лучшие сорта этой культуры дают высокий урожай зеленой массы и зерна, поэтому у нас в стране и за рубежом ведется интенсивная работа по созданию сортов тритикале, приспособленных к почвенно-климатическим условиям соответствующей зоны.
Октаплоидные тритикале получают при скрещивании с рожью мягкой пшеницы, гексаплоидные — твердой и других видов пшениц, имеющих 2n=28 хромосом (схема).

Путем аллоплоидий можно получать растения, содержащие геномы разных видов и родов, и создавать новые формы, не существующие в природе. Этим методом можно воссоздавать (ресинтезировать) уже существующие виды, предки которых исчезли. в 1930 г, с помощью аллополиплоидии осуществил ресинтез культурной сливы Он скрестил тёрн с алычой. Среди гибридных растений ему удалось выделить одно, сходное с культурной сливой, в клетках которого содержались присущие этому виду 48 хромосом:

Гетероплоидами или анецплоидами называют организмы, число хромосом у которых некратное гаплоидному Гетероплоиды могут возникать разными путями. В одних случаях они образуются в результате отхождения двух гомологичных хромосом к одному полюсу в анафазе I мейоза или в анафазе митоза. Чаще всего они образуются в результате отсутствия конъюгации гомологичных хромосом и образования унивалентов. Униваленты, как правило, не ориентируются надлежащим образом и могут отойти к одному полюсу. Причиной возникновения гетероплоидов может быть также отсутствие разделения хромосом на хроматиды. В этом случае нарушается их расхождение в дочерние клетки при втором делении мейоза. В том и другом случаях могут образоваться гаметы с набором хромосом, некратным гаплоидному: п—1, n+1. При слиянии этих гамет в процессе оплодотворения могут образоваться зиготы, содержащие 2n— 1, 2n—2, 2п+1 и 2n+2 хромосом. В зависимости от числа дополнительных или недостающих хромосом применяют следующие термины: 2п—112— моносомик, 2п—212 — нуллисомик, 2п+15 — трисомик, 2n+25 — тетрасомик. Цифра внизу указывает номер хромосомной пары в кариотипе, в которой изменилось число хромосом.
Гетероплоидию в генетике растений попользуют для определения групп сцепления генов, в селекции — для получения межсортовых замещенных линий и создания так называемых дополненных линий, одна пара хромосом у 'которых замещена идентичной парой гомологичных хромосом другого сорта, в которой содержатся гены, контролирующие хозяйственно ценные признаки. Для этого создается полный набор гетероплоидных форм по всем парам гомологичиых хромосом. Впервые полные серии мносомиков и нуллисомиков были получены Э. Сирсом у мягкой яровой пшеницы сорта Чайниз Спринт (Китайская яровая) в 40—50-х годах. Нуллисомики по разным парам гомологичных хромосом четко различаются по высоте растений, (морфологическому отроению, величине и стерильности колоса, поэтому можно определить локализацию генов в соответствующих хромосомах.
Полиплоидия у животных и человека. Полиплоидия у животных встречается крайне редко. Единственный известный случай полиплоидии у млекопитающих — золотистый хомячок, в кариотипе которого содержится 44 хромосомы, в то время как у животных других родов серого и обыкновенного хомяка их 22. Искусственно тетраплоидные формы удавалось получать у некоторых видов рыб и амфибий, но сохранить тетраплоидное число хромосом в потомстве и даже просто получить потомков не удавалось. Так, у аксолотля были получены тетраплоидные самки. При скрещивании их с диплоидными самцами было получено триплоидное, полностью бесплодное потомство. При скрещивании двух подвидов японской лягушки были получены аллоплоиды, но они были бесплодны.
Отмечен единичный случай рождений мальчика-триплоида, в генотипе которого содержалось 66 аутосом и XXY-половых хромосом; масса при рождении 2190 г; видимых нарушений в развитии отдельных частей тела не наблюдалось. Среди абортированных плодов человека отмечены также случаи образования триплоидных эмбрионов.
Гетероплоидия наиболее изучена у человека. Установлено, что хромосомные нарушения определяют мертворождение или смерть новорожденных в течение первого и последующих лет жизни. Вместе с тем в некоторых случаях рождаются и живут относительно продолжительное время дети-трисомики по какой-либо хромосоме, но во всех случаях трисомия вызывает пороки развития. В 1960 г. был описан синдром Патау — тяжелое заболевание, обусловленное трисомией по 13-й хромосоме. Частота встречаемости — 1 : 5000—7000 новорожденных. При этом наблюдаются высокая ранняя смертность, пороки головного мозга и лица, полидактилия (многопалость), пороки внутренних органов, в том числе перегородок сердца.
Синдром, или болезнь, Дауна обусловлен трисомией по 21-й хромосоме, наиболее часто встречается у новорожденных, в среднем составляет около 1 на 700—800 рождений. Трисомия по этой хромосоме бывает причиной ряда пороков развития: пороки сердца, пищеварительного тракта, патология в форме головы и лица, разболтанность суставов, умственная отсталость.
Причиной трисомии является неравное расхождение хромосом в мейозе у одного из родителей, чаще — у матери Причин нарушения мейоза может «быть много, в том числе и возраст матери, как это установлено для синдрома Дауна.
Весьма разнообразны случаи гетероплоидии у человека по половым хромосомам. По данным , частота их встречаемости около 1,6:1000 рождений. Моносомия по Х-хромосоме обусловливает синдром Шерешевского—Тернера. Для него характерны бесплодие, недоразвитие половых признаков, врожденные соматические пороки развития, низкий рост.
Довольно часто встречается трисомия по половым хромосомам у мальчиков. Причем, если в кариотипе присутствует несколько дополнительных Х-хромосом и хотя бы одна У-хромосома, рождаются мальчики. Частота таких рождений составляет 1,39—1,98 на 1000 рождений мальчиков. Отмечены случаи рождения мальчиков и с ди - и трисомией по У-хромосоме (синдром Клайнфельтера). В этом случае в начальный период развития у больных не наблюдается существенных аномалий, но для них, как правило, характерно бесплодие.
Хромосомные аберрации (перестройки). Изменение структуры хромосом вследствие их разрывов и перестроек называют хромосомными аберрациями. Любому структурному изменению хромосомы предшествует ее разрыв, при котором получаются два фрагмента, каждый из них имеет по одному «клейкому» концу, а они, в свою очередь, способны соединиться с любым другим «клейким» концом этой или другой хромосомы. Характер хромосомной перестройки во многом зависит от состояния хромосомы в момент воздействия мутагенного фактора. Если хромосома находится в состоянии одиночной нити (период G\ интерфазы, анафаза и телофаза митоза), то в последующий период S интерфазы она удваивается и аберрация сохраняется в обеих хроматидах, то есть возникают хромосомные аберрации.
Если мутаген действует на хромосому, находящуюся в состоянии двойной нити (период G2 или S интерфазы, профаза и метафаза митоза), аберрация может произойти только в одной хроматиде. В этом случае возникают хроматидные перестройки.
Различают внутри - и межхромосомные аберрации. К внутри-хромосомным относят делеции, дефишенси, инверсии, дупликации и фрагментации. К межхромосомным — транслокации. Тип хромосомной аберрации обозначают символом с указанием порядкового номера хромосомы, в которой она произошла. Например, если в 5-й хромосоме произошла делеция, ее обозначают символом Dl (5), инверсия In (5), транслокация между 5-й и 1-й хромосомами обозначается Т (5—1) (рис. 49).
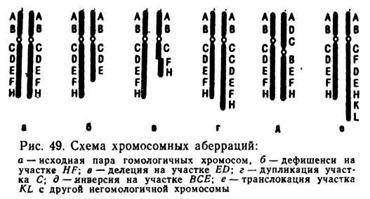
Делеция — выпадение участка хромосомы в средней ее части, содержащего обычно целый комплекс генов. В случае выпадения концевого участка возникает концевая делеция — дефишенси. Когда делеция и дефишенси захватывают небольшой фрагмент хромосомы, это вызывает изменение признака, например желтую окраску тела и белоглазие у дрозофилы. Крупные делеции, как правило, легальны и вызывают гибель организма. Известна крупная делеция 21-й хромосомы человека, которая вызывает тяжелую форму белокровия.
Инверсия (In) возникает в результате разрыва хромосомы одновременно в двух местах с сохранением внутреннего участка, который воссоединяется с этой же хромосомой после поворота на 180°. В этом случае группа сцепления генов в данной хромосоме сохраняется, но изменяется положение генов относительно друг друга. Инверсия не влияет на фенотип особи, но при этом нарушается конъюгация гомологичных хромосом в мейозе и в анафазе I образуются инверсионные мосты.
Дупликация (Dp) — удвоение участка хромосомы. Обычно дупликация не оказывает сильного влияния на фенотип особи. Вместе с тем увеличение дозы одного и того же гена может вызвать фенотипическое изменение характера проявления признака, как это имеет место у дрозофилы при дупликации гена Ваr (полосковидные глаза). При дупликации данного гена уменьшается число фасеток в глазах насекомого и усиливается деформация глаз.
Фрагментация (F) происходит в результате разрыва хромосом или хроматид в нескольких местах одновременно и образования отдельных фрагментов хромосом с последующей утерей в митозе тех из них, которые не содержат центромеры. Как правило, фрагментация обусловливает возникновение летальных мутантов.
Транслокация (Т) — обмен участками между негомологичными хромосомами; ее относят к межхромосомным аберрациям, так как структурные изменения происходят одновременно в двух или более негомологичных хромосомах. Транслокации не изменяют числа генов в данном генотипе и не всегда проявляются фенотипически, но у особей, гетерозиготных по транслокации, нарушается конъюгация гомологичных хромосом и образуются нежизнеспособные гаметы. Японский ученый Й. Тазима в 1959 г, при обработке икс-лучами тутового шелкопряда перенес на Y-хромосому (половая хромосома у тутового шелкопряда детерминирует женский пол) ген черной окраски грены. X-хромосома не несет этого гена, поэтому белые яички дают самцов, черные — самок. С помощью фотоэлемента можно осуществить сортировку грены и снабжать шелководов греной, из которой выводятся только самцы, образующие более крупные коконы.
Генные мутации. Генными, или точковыми, мутациями называют изменения структуры молекулы ДНК на участке определенного гена, кодирующего синтез соответствующей белковой молекулы. Следует отметить, что молекула ДНК проявляет относительно высокую стабильность и устойчивость к мутагенам, обладает свойством восстанавливать первоначальную структуру и исправлять повреждения, если они затрагивают только одну из комплементарных цепочек. Процесс восстановления первоначальной структуры и исправления повреждений молекулы ДНК называется репарацией. Наиболее изучены фотореактивация и темновая репарация,
Фотореактивация осуществляется фотореактивирующим ферментом. Свет активирует фермент, и он восстанавливает исходную структуру молекулы ДНК, поврежденную ультрафиолетовыми лучами.
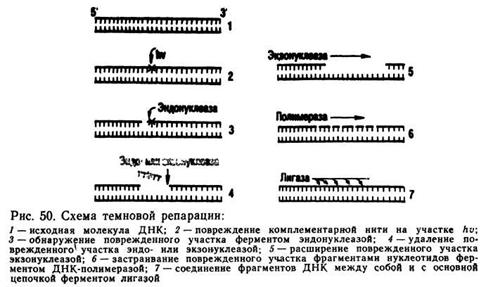
Темновая репарация — механизм исправления различных повреждений молекулы ДНК, вызванных химическими или физическими мутагенами. Темновая репарация протекает в несколько этапов, которые были установлены на примере Е. coli P. Сетлоу в 1964 г. При этом участвуют четыре типа ферментов, последовательное действие которых исправляет повреждение ДНК, если на данном участке повреждена только одна из двух комплементарных нитей ДНК. Схематично этот процесс может быть представлен следующим образом (рис. 50):
1. Фермент эндонуклеаза «обследует» молекулу ДНК, опо
знает место повреждения, вблизи него «надрезает» нить ДНК в
начале и в конце поврежденного участка и удаляет его.
2. Фермент экзонуклеаза расширяет поврежденный участок,
удаляя из нити ДНК 500—1000 нуклеотидов, примыкающих к
поврежденному участку. Такое расширение места повреждения
необходимо для последующего его «застраивания».
3. Фермент ДНК-полимераза синтезирует удаленный участок
молекулы ДНК, располагая нуклеотиды комплементарно второй
неповрежденной нити.
4. Фермент лигаза скрепляет синтезированные ДНК-полимеразой фрагменты ДНК друг с другом и с концами поврежденной нити ДНК. Таким образом, осуществляется полное восстановление поврежденных участков молекулы ДНК, и она приобретает первоначальную структуру.
Репарация молекулы ДНК, как правило, протекает в период G1 митотического цикла. При этом происходит исправление структурных повреждений молекулы ДНК, разрывов полинуклеотидных цепей, удаление некомплементарных нуклеотидов.
Если в молекуле ДНК на одном и том же участке одновременно повреждаются обе комплементарные нити, то это повреждение не восстанавливается и проявляется в виде генных, или точковых, мутаций. Мутации могут возникать в результате выпадения или вставки нуклеотидных пар в молекуле ДНК на участке соответствующего гена или замены одного нуклеотида на другой, когда вместо тимина становится гуанин или вместо гуанина — аденин.
Под действием мутагена во втором триплете выпадает нуклеотид тимин, а в седьмом произошла вставка нуклеотида тимина. Вследствие этого на участке данного гена произойдет «сдвиг рамки считывания» мРНК и в полипептидной цепи будут кодироваться другие аминокислоты:
![]()
В том случае, когда произошло выпадение или добавление хотя бы одной пары нуклеотидов, нарушается транскрипция — сдвигается «рамка считывания» при синтезе мРНК, изменяется порядок чередования аминокислот в полипептидной цепи, кодируемой данным геном. Так, например, на одном из участков ДНК у вируса Т4 в норме имеется следующее чередование триплетов, обеспечивающее соответствующий порядок расположения аминокислот в полипептидной цепи:

Фенотипическое проявление мутации зависит от того, на каком участке произошла вставка или выпадение нуклеотидной пары. Если она выпала вблизи промотора, то есть в начале структурного гена, то транскрибируется сильно измененная мРНК, транслируется «испорченная» полипептидная цепь и белковая молекула, не выполняя своей функции, быстро инактивируется. Если вставка или выпадение нуклеотидной пары произошли на конечном участке данного гена в молекуле ДНК или, как в нашем примере, выпадение сочетается с добавлением нуклеотидной пары, то это не приведет к инактивации белковой молекулы, но повлияет на качество кодируемого белка и обусловит изменение признака или свойства.
При замене в триплете ДНК одного нуклеотида другим в полипептидной цепи произойдет замена только одной аминокислоты. Так, например, на участке ДНК, кодирующем белок глобин человека, в триплете ГТЦ нуклеотид гуанин может замениться тимином и в полипептидной цепи вместо аминокислоты глутамина будет кодироваться лизин. Такое явление получило название «миссенс-мутация».
Миссенс-мутацией называют замену на участке структурного гена одной нуклеотидной пары другой, в результате чего кодируется включение в полипептидную цепь «неправильной» аминокислоты. В этом случае в молекуле ДНК возникает новая аллель данного гена, происходит мутационное изменение фено-тйпического проявления признака. Это явление получило название множественного аллелизма.
Множественным аллелизмом называют различное состояние одного и того же гена (локуса), обусловленное точковыми мутациями, детерминирующими различное проявление одного и того же признака или свойства. Аллели одного гена, возникшие в результате точковой мутации, называют множественными аллелями.
Впервые множественный аллелизм был установлен в 1929— 1930 гг. , и вым у дрозофилы на примере локуса гена scute. Множественные аллели этого гена — sc1, sc2, sc3 — вызывали различный характер редукции щетинок на теле дрозофилы.
Ярким примером множественного аллелизма могут служить аллели, кодирующие синтез глобина — белка, необходимого для образования сложных молекул гемоглобина крови. Замена только одной из 300 аминокислот в белковой молекуле глобина обусловливает новый тип гемоглобина. В настоящее время известно около 100 типов гемоглобина, контролируемых серией множественных аллелей. Например, молекула глобина А (нормальный тип гемоглобина) на одном из участков полипептидной цепи имеет следующий порядок чередования аминокислот: — пролин — глутаминовая кислота — глутаминовая кислота — лизин —. Замена нуклеотидов на данном участке ДНК обусловливает кодирование другого типа гемоглобина:

В гомозиготном состоянии гемоглобин обусловливает тяжелое наследственное заболевание — серповидно-клеточную анемию.
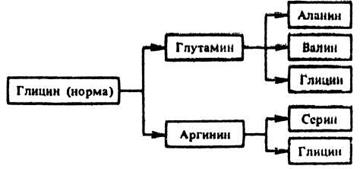
Примером множественного аллелизма является нарушение кодирования фермейта триптофансинтетазы, обеспечивающего синтез аминокислоты триптофана у кищечной палочки E. coli,
На участке гена, кодирующего синтез данного фермента, вследствие точковой мутации происходит изменение порядка чередования нуклеотидов в молекуле ДНК на участке триплета, кодирующего аминокислоту глицин. Это определяет замену глицина в полипептидной цепи глутамином или аргинином.
Интересно, что у животных белковые молекулы, выполняющие сходные функции, различаются небольшим числом аминокислот. Так, молекулы инсулина у животных разных видов отличаются составом аминокислот только на одном участке молекулы (табл. 4).

Генные мутации могут возникать не только в одном, но и в разных генных локусах, имеющих сходное влияние на характер развития признака, поэтому при работе с мутантами бывает необходимо установить, действительно ли причиной мутации являются множественные аллели. Для этого используют метод, предложенный и получивший название «критерий аллелизма. Если при скрещивании двух мутантов в F1 проявляется признак одного из них, a a F2 наблюдается расщепление в отношении 3: 1, то имеет место множественный аллелизм одного гена.
Если при скрещивании двух мутантов в F1 проявляется признак дикого типа, а в F2 имеет место расщепление в отношении 9:7, как при комплементарном взаимодействии, то мутировали разные гены:

Множественные аллели обозначают символом основного гена с буквенным либо цифровым знаком. Например, у томатов в локусе длинного плеча второй хромосомы известна серия аллелей, детерминирующих различную высоту растений. Аллель d+ (дикий тип) контролирует нормальную высоту растений, он доминантен к другим аллелям данной серии. По степени доминирования при выращивании растений в теплице аллели располагаются следующим образом:
![]()
Серией множественных аллелей детерминируется и полиморфизм белков у животных. В 1957 г. Эштон у крупного рогатого скота установил полиморфизм белков трансферрина, составляющего около 3—6% сыворотки крови. Белок трансферрин состоит из одной полипептидной цепи, включающей около 750 аминокислот. Полиморфизм трансферрина проявляется в виде различных фракций при электрофорезе. Локус трансферрина обозначают буквами Tf, а его аллели — TfA, TfD, TfE. Наследуются они по типу кодоминирования, то есть в гетерозиготном состоянии проявляются фракции, кодируемые обоими аллелями.
Наследование признаков, контролируемых множественными аллелями, имеет сложный характер. Различают следующие основные типы доминирования:
полное доминирование; доминантный аллель подавляет проявление любого другого аллеля данной серии. Остальные аллели детерминируют более слабое проявление признака, а в гетерозиготном состоянии обладают кумулятивным (суммирующим) эффектом. По такому типу наследуется синтез пигмента меланина, определяющего интенсивность окраски меха у гвинейской (морской) свинки. Если принять количество меланина, синтезируемого животными, имеющими генотип СС, за 100%, а саса— за 0 (альбиносы), аллель сr —42%, a cd — 39%, то при полном доминировании аллеля С у животных, имеющих генотипы СС, Ccr, Ccd, Cca, будет содержаться 100% меланина и они будут иметь темную окраску. Содержание меланина у животных» имеющих другие генотипы, зависит от сочетания аллелей: животные с генотипом сrсr будут иметь 84% меланина, crcd— 74, сrса — 42, cdcd— 67, cdca — 39%, саса — 0, то есть будут альбиносами;

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
