
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Организмам разных видов присущ врожденный иммунитет. Примерами врожденного иммунитета служит невосприимчивость крупного рогатого скота к сапу, которым тяжело болеют лошади; алжирские овцы не болеют сибирской язвой; куры породы белый леггорн более устойчивы к пуллорозу, чем куры пород род-айланд, плимутрок, виандот. Высокой устойчивостью среди птиц обладают цесарки. Известно, что разные породы собак проявляют разную чувствительность к вирусу чумы. Более чувствительны и тяжело переносят все три типа чумы (легочную, кишечную, нервной ткани) немецкая и южнорусская овчарки и лайки, а фокстерьер и эрдельтерьер более устойчивы и при заболевании переносят чуму легче и без серьезных последствий,
Следовательно, породная, индивидуальная естественная резистентность и иммунитет могут быть результатом естественного отбора или направленной селекции в целях формирования наследственно устойчивого иммунитета.
Естественная резистентность. Кожный покров, слизистые оболочки дыхательных путей, кишечника, мочеполовой системы и клетки этих тканей защищают организм путем выделения защитных биохимических веществ. Важную функцию в защите организма выполняет кровь. В обеспечении защиты велика роль костной ткани, тимуса, селезенки, печени, лимфатической системы, клетки которых служат источниками иммунной защиты.
Иммунную роль выполняют различные клетки: Т-лимфоциты тимуса, В-лимфоциты костного мозга, макрофаги, нейтро-фильные гранулоциты, базофилы, эозинофилы и другие типы иммуноцитов.
Основные процессы формирования лимфоцитов определены функцией ряда органов:
печень плода, в которой образуются стволовые кроветворные клетки - предшественники лимфоцитов;
костный мозг — источник В-лимфоцитов. Здесь формируются стволовые клетки, которые покидают костный мозг в период эмбриогенеза и переходят в лимфоидные органы;
тимус, в котором формируются Т-лимфоциты; им принадлежит ведущая роль в становлении клеточного иммунитета, так как они стимулируют синтез антител В-лимфоцигами. Клетки, покидающие тимус, превращаются в лимфоциты и, переходя в кровь, обладают полной иммунокомпетентностью. Тимус увеличивается в размерах до периода половой зрелости, а в процессе старения происходит его физиологическая инволюция;
лимфатические узлы; это вторичные лимфоидные органы, которые улавливают антигены из лимфы. В ткани лимфоузла встречаются лимфоциты и макрофаги. Под влиянием проникающих в узел антигенов из лимфоцитов развиваются иммуно-бласты, из которых образуются иммунокомпетентные лимфоциты, участвующие в клеточном иммунитете;
селезенка; выполняет роль фильтра для крови, удаляет отработанные клетки крови и образует новые лимфоциты. У птиц первичным лимфоидным органом является фабрициева сумка (бурса).
Неспецифический иммунитет включает гуморальные и клеточные факторы.
Гуморальные факторы. К этой группе относят бактерицидную активность крови (БА), которая объединяет антимикробную активность таких веществ, как комплемент, пропердин, нормальные антитела, лизоцим, бета-лизин, и действует на оболочку бактерии, которая разрушается лизоцимом.
Лизоцим — это фермент, молекула которого имеет одну цепочку из 129 аминокислотных остатков. Лизоцим синтезируется в клетках макрофагов и локализуется в лизосомах. Он широко распространен во всех биологических жидкостях: в околоплодной жидкости, молозиве, секретах слизистых оболочек, сыворотке крови, Его защитная роль обусловлена способностью лизировать оболочку многих видов микробов, особенно из грам-положительной микрофлоры. Ферментативная активность лизоцима в большей степени связана с третичной структурой его молекулы. Таким образом, лизоцим является клеточным фактором неспецифической защиты, а его концентрация в исследуемой жидкости (ткани) служит показателем фагоцитарной активности организма. Уровень лизоцимной активности обусловлен наследственностью и может подвергаться селекционному воздействию, повышаясь у животных в различных линиях, семействах, у потомства разных производителей в результате селекционного подбора в породе или стаде.
Бета-лизин — это пептид, содержащий большое количество лизина и синтезируемый тромбоцитами. Он обнаружен в сыворотке крови, слюне, плазме, в легких, кишечнике, печени и др. Бета-лизин воздействует на грамположительные микроорганизмы, точнее — на цитоплазматическую мембрану бактерий, и вызывает их лизис,
Комплемент является сложным белком ферментного типа и состоит из 9 компонентов разного ферментного действия. У птиц в комплемент входят 4 компонента; у лошадей, крупного рогатого скота — 9 компонентов. Комплемент способен соединяться с комплексом антиген — антитело. Он разрушает липидную оболочку микроба под действием эстеразного комплекса и способствует процессу фагоцитоза. Синтез комплемента происходит в клетках тонкого отдела кишечника, лимфоцитах, клетках селезенки, лимфоузлов, костного мозга.
Пропердин играет важную роль в естественной неспецифической резистентности. В его состав входят белок сыворотки крови, ионы магния и комплемент. Действие его осуществляется совместно с другими защитными факторами.
Интерферон — неспецифическое противовирусное вещество, синтезируемое лейкоцитами; действует на разные вирусы и быстро образуется в клетке с появлением вируса, влияя на нуклеиновые кислоты в период репликации их РНК.
Холинэстераза — это высокоактивный фермент. Он может служить показателем неспецифической резистентности организма. Присутствует в плазме крови, препятствует распространению ацетилхолина в тканях.
К группе гуморальных факторов можно отнести естественные антитела, которые присутствуют в крови или иной жидкости. Естественные антитела синтезируются в В-лимфоцитах, они усиливают клеточную защиту в виде фагоцитоза, стимулируют функцию рецепторов В - и Т-клеток, способствуют скучиванию и разрушению микробных клеток и нейтрализации токсинов. Антитела разделяют на сывороточные (в крови, молозиве, молоке) и на секреторные (на поверхности слизистых оболочек).
Неспецифическая гуморальная защита проявляется в виде так называемого бактерицидного эффекта (или активности), которая обеспечивается действием пропердина, лизоцима, комплемента, интерферона, бета-лизина, антител. Бактерицидная активность служит интегральным показателем гуморальных факторов естественной резистентности.
Клеточный тип защиты. Кроме гуморального иммунитета, важную роль в защите организма играет клеточный иммунитет, особенно необходимый при некоторых инфекциях, отторжении трансплантантов при пересадке органов или эмбрионов, аутоиммунных заболеваниях (в этом процессе клеточный иммунитет является формой защиты организма от инфекций, вызванных микроорганизмами).
Клеточный тип защиты обусловлен фагоцитозом, который осуществляют многие клетки: микрофаги (гранулярные лейкоциты и лимфоциты), макрофаги (ретикулоэндотелий печени, селезенки, костного мозга, лимфатических желез), подвижные клетки соединительной ткани (гистоциты, моноциты). Наиболее активно фагоцитируют полиморфно-ядерные зернистые лейкоциты. Тромбоциты, выделяя агглютинин, повышают активность нейтрофильных гранулоцитов.
Основным элементом иммунной системы служат популяции лимфоцитов двух основных типов: лимфоциты типа В и Т, символы которых приняты в 1969 г.
В-лимфоциты формируются в костном мозге. Их основная функция состоит в синтезе антител, то есть иммуноглобулинов, которые служат источником гуморальных факторов иммунитета.
Т-лимфоццты образуются в тимусе. Они не вырабатывают антитела, а выполняют защитную роль с помощью рецепторов, находящихся на поверхности лимфоцита.
Рецепторы — это образования макромолекулярной структуры В - и Т-лимфоцитов, расположенные на поверхности этих клеток и обеспечивающие «узнавание» конкретного антигена. Рецепторные клетки имеют специфичность, обусловленную генетически в процессе их биосинтеза, что создает возможность распознавания каждого антигена.
Т-лимфоциты могут выполнять разные функции. Существуют Т-киллеры, которые, соединяясь с чужими клетками, убивают их. Другие Т-лимфоциты содействуют В-лимфоцитам в синтезе антител. Третий тип Т-лимфоцитов — это Т-супрессоры, подавляющие функции В-клеток,
Фагоцитоз регулируется нервной системой с помощью медиаторов, выделяемых нервными окончаниями. Процессу фагоцитоза способствует активация макрофагов антителами (опсонинами) и комплементом.
Иммунитет организма обеспечивает защиту не только от микроорганизмов, но и от других генетически чужеродных веществ, особенно при пересадках органов и тканей; от собственных переродившихся клеток при изменении их генетического аппарата, в том числе раковых.
Регуляция иммунного ответа осуществляется путем специфической стимуляции лимфоцитов, что приводит либо к биосинтезу антител, либо к усилению клеточного иммунитета. На регуляцию иммунного ответа оказывают влияние некоторые гормоны, например катехоламин и гормоны надпочечников. Кортикостероиды влияют на созревание, дифференцировку и распределение лимфоцитов.
Иммунная система характеризуется тесным взаимодействием факторов гуморального и клеточного иммунитета. Это проявляется в том, что гуморальные вещества, такие как опсонины, агглютинины и другие антитела, синтезируемые иммуно-компетентными лейкоцитами, способствуют осуществлению клеточного иммунитета, то есть реакции фагоцитоза. Фагоцитарные реакции не могут уничтожать вирусы и токсины, на которые активно действуют гуморальные факторы. Вместе с тем клеточная реакция способствует образованию антител, нейтрализующих токсины. Следовательно, гуморальные и клеточные факторы проявляют единство в системе защиты организма.
Онтогенетические этапы образования лимфоцитов. В периферической крови циркулируют стволовые клетки, которые являются предшественниками лимфоцитов. В конце эмбрионального периода органом кроветворения является печень, а затем костный мозг. В первые дни после рождения в циркулирующей крови в десятки раз больше стволовых клеток, чем у взрослых животных.
Выброс стволовых клеток из костного мозга в периферическую кровь имеет суточную цикличность и связан с обратным направлением суточного выброса в кровь гормонов кортикостероидов. Следовательно, поступление в периферическую кровь лимфоцитов находится под гормональным контролем со стороны коры надпочечников и гипофиз адреналиновой системы.
Существуют популяции лимфоидных клеток, обладающих способностью сохранять информацию о каком-либо антигене в течение длительного периода времени («клетки памяти»). Это создает условия сохранения длительного иммунитета к данному антигену, однако «иммунная память» лимфоцитов не наследуется.
Установлено (Петров, 1966), что существуют генетически сильные и слабые продуценты антител. Эти данные подтверждают возможность селекционной работы с животными в целях повышения естественной резистентности в поколениях.
(1984) разработана схема онтогенетических изменений, происходящих в иммунной системе (рис. 62).
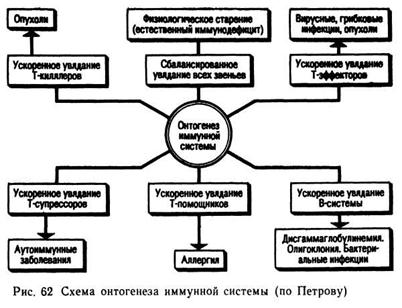
На формирование активности иммунной системы и резистентности большое влияние оказывают возрастные процессы, так как с ними связаны не только интенсивность физиологического состояния организма, но и накопление в клетках и тканях организма различных мутационных эффектов в виде новых генов, которые являются источником новых антигенов, образования опухолей, патологии различных процессов, что и отражено в приведенной схеме.
В процессе старения происходит постепенное уменьшение числа стволовых клеток костного мозга, снижается фагоцитарная активность, сокращается синтез антител, уменьшается миграция клеток костного мозга в кровь, наступает снижение общей иммунной активности и появление аутоиммунных антител как результат патологии.
Вопрос о продлении иммунной активности, уменьшении процессов возрастной иммунной депрессии имеет важное значение для практики племенного животноводства, когда для селекционных целей целесообразно продлевать сроки использования ценных производителей и самок.
Иммунореактивность. Способность иммунной системы Своевременно отвечать на проникновение инфекции называют иммунореактивностью. Реакция зависит от концентрации антител (иммуноглобулинов) и соотношения численности и связи между Т - и В-лимфоцитами.
Иммунореактивность зависит и от гормонального фона, например от кортикостероидов. На уровень реактивности оказывают влияние биоритмы. Факторами, нарушающими иммунную систему и иммунореактивность организма, являются различного типа стрессы, неблагоприятные для организма факторы среды, которые нарушают необходимый иммунный гомеостаз.
Иммунные реакции организма на воздействия могут отклоняться от нормального ответа и проявлять иные, специфические реакции на антигены, чаще всего имеющие патологический характер. Так, например, известна повышенная чувствительность организма к разным антигенам, проявляющаяся в виде аллергии, которая часто протекает в острой патологической форме заболевания. Гиперчувствительность может проявляться в виде анафилаксии и анафилактического шока, которые вызывают опасные симптомы удушья, падения кровяного давления и др.
В противоположность перечисленным реакциям существует явление иммунологической толерантности* к антигенным факторам. Это, в частности, проявляется в отсутствии иммунного ответа материнского организма на антигены плода во время беременности, что спасает плод от его отторжения.
Существует и такая форма иммунного ответа, когда организм начинает синтезировать антитела на антигены собственного организма (аутоантитела), например к гормонам щитовидной железы, что приводит к серьезным нарушениям в обмене веществ. У животных появление аутоантител может происходить в отношении своих гамет, что приводит к бесплодию.
В последнее десятилетие выявлен так называемый синдром приобретенного иммунодефицита (СПИД), при котором иммунная система организма утрачивает свою защитную функцию под влиянием вируса иммунодефицита человека (ВИЧ) и это приводит к неизбежной гибели людей. Борьба со СПИДом приобрела глобальный характер.
Генетическая обусловленность иммунной системы. Основная способность иммунной системы заключается в определении «своего» и «чужого» в организме. Ее реакция на «чужое» сводится к устранению его из организма.
Вещества, вызывающие иммунный ответ организма на «чужое», называют антигенами. Ими могут быть различные белки, полисахариды, микроорганизмы (бактерии и их токсины, вирусы, грибы), аллергены (выделения растений или насекомых, комнатная пыль), антигены групп крови, трансплантационные антигены пересаживаемых тканей, чужеродные вещества других организмов.
Свойство антигенов вызывать иммунную реакцию организма называется иммуногенностью. Каждый антиген вызывает специфическую реакцию организма в виде синтеза специфических веществ (антител), образуемых в организме, подвергнутом воздействию антигенов. Не имеют антигенных свойств нуклеиновые кислоты, липиды (жиры, воск).
Антигенными свойствами обладают эритроциты. Набор антигенов у эритроцитов имеет специфичность и индивидуальность у каждого организма. Эта индивидуальность, то есть присущие организму группы крови, должна учитываться, если необходимо перелить кровь донора в организм реципиента. Если эритроцитарные антигены донора и реципиента несовместимы, то переливание крови проводить нельзя, иначе произойдут патологические процессы и даже гибель реципиента.
Реакция антиген — антитело специфична, что объясняется генетической специфичностью антител, соответствующих определенному антигену. Реакция антиген — антитело может проявляться в виде агглютинации, преципитации, лизиса и др. Эти реакции используют для диагностики протекающего иммунного ответа организма.
Генетические и физиологические особенности иммуноглобулинов. Основным элементом иммунной системы являются антитела, синтезируемые иммунокомпетентными лимфоцитами в качестве реакции на антиген. С 1964 г. антитела принято называть иммуноглобулинами (Ig). Выявлено пять различных классов: IgG, IgM, IgD, IgE, IgA. Иммуноглобулины представляют собой белки сыворотки крови. В реакции антиген — антитело иммуноглобулины реагируют только с определенными антигенами; это обусловлено специфической аминокислотной последовательностью N-концевого участка их молекулы, что является результатом строения ДНК клеток, синтезирующих иммуноглобулины.
Иммуноглобулин G обнаружен в сыворотке крови и молозиве жвачных. Он составляет около 75% от общего количества иммуноглобулинов. IgA составляет около 10% всех иммуноглобулинов сыворотки крови; он найден в большом количестве в пищеварительной системе и в молозиве.
Структура молекулы иммуноглобулинов независимо от типа иммунного ответа имеет ряд общих элементов. В нее входят две тяжелые полипептидные цепи Н (heary) и две легкие L (tight), которые короче цепей H. Эти полипептидные цепи соединяются между собой дисульфидными связями. Тяжелые H-цепи обладают антигенностью, и на них организм отвечает синтезом антител. В состав H-цепей входит 420 — 440 аминокислотных остатков. H-цепи входят в состав существующих 5 классов иммуноглобулинов. Тяжелые цепи имеют обозначение для каждого класса иммуноглобулинов, а именно: γ для IgG, μ для IgM, α для IgA, δ для IgD, ε для IgE.
Концевая часть H - и L-цепей, несущая NHз-остатки, характеризуется большой изменчивостью по составу входящих аминокислот и обозначается символом V (variable). Эта вариабельная часть молекулы Ig обеспечивается генами локуса V, который определяет порядок размещения 97 аминокислот L-цепй, Противоположный конец молекулы имеет постоянный состав по аминокислотам и обозначен буквой С (constant). Цепи L и H различаются не только по длине, но и по составу. Так, цепи L имеют четыре формы: λ1, λ2, λ3 и x — состоящие из 210 — 230 аминокислотных остатков. Эти цепи присущи всем пяти классам иммуноглобулинов,
Генетическая особенность и изменчивость молекул иммуноглобулинов обусловлена структурой N-концевой части H- и L-цепей.
Другой концевой (С) участок цепей отличается постоянством состава цепи по аминокислотам, V- и С-участки цепей ДНК кодируются разными генами трех хромосом. У мыши локусы H-цепи находятся в 12-й хромосоме; локус х L-цепи — в 6-й хромосоме, а локус λ L-цепи — в 16-й хромосоме.
Причиной большого разнообразия антител является соматическое мутирование, разрывы и рекомбинации между локусами V- и С-генов, Следовательно, специфичность и разнообразие антител в иммунной системе обусловлены особенностями в структуре и функциях V- и С-частей тяжелых и легких цепей молекулы иммуноглобулина.
Биосинтез и генетика антител. В основе синтеза антител на первом этапе этого процесса лежит матричный механизм, обусловленный генной структурой ДНК и РНК ядер иммунокомпетентных лимфоцитов и генетическими особенностями ферментов.
Синтез H- и L-цепей Ig детерминирован одним С-геном, а разнообразие цепей определяется большим числом Y-генов, Каждый из двух С-генов H- и L-цепей сочетается с Y-геном и образует структуру, называемую цистроном. Цистрон является исходным генетическим элементом. Его генетическая информация копируется однонитевой молекулой РНК, которая приобретает информационную генетическую функцию, преобразуясь в иРНК.
Информационная РНК переходит из ядра лимфоцита в его цитоплазму и на полирибосомы цитоплазмы. Начинается синтез полипептидных цепей. H- и L-цепи синтезируются на разных рибосомах. Продолжительность этого синтеза — около 30—90 с. После образования H- и L-полипептидные цепи соединяются дисульфидными мостиками и происходит окончательная сборка молекулы иммуноглобулина, включающая четырехцепочечную структуру. Образуется специфическое антитело, синтез которого был ответом организма на воздействие чужеродного антигена.
Скорость синтеза молекул Ig велика. Так, внутри одной клетки за одну секунду синтезируется около двух тысяч одинаковых молекул.
Установлено, что особи одного вида различаются по скорости образования антител на один и тот же антиген. Такие различия в скорости реакции биохимического процесса в иммуно-компетентных В - и Т-клетках обусловлены видовыми и индивидуальными генетическими особенностями каждой клетки. Это означает, что существует локус, определяющий степень экспрессии гена в синтезе Ig, Этот локус назвали локусом иммунного ответа, который обозначен символом Ir.
Классы иммуноглобулинов Иммуноглобулины различаются по составу H-цепей и образуют пять классов, каждый из которых имеет физико-химическую особенность и биологическую активность
Легкие цепи входят в состав всех пяти классов иммуноглобулинов. IgG наиболее распространен. Он обладает способностью проходить через плаценту и тем самым защищает развивающийся зародыш. Период полужизни IgG —21 день. Он более активен в присутствии комплемента и опсонореакции. IgM в процессе эволюции был филогенетически более ранним. В онтогенезе он также синтезируется раньше других иммуноглобулинов (у плода и новорожденного синтезируется в основном IgM), IgM находится преимущественно в плазме крови и в лимфе, период полужизни — 5,1 дня. Активируется в реакции агглютинации и лизиса в присутствии комплемента. Он не обладает способностью проникновения через плаценту. При инфекциях уровень IgM повышен. IgA содержится в сыворотке крови, слюне; типичен для слизистых оболочек и осуществляет местную защиту от инфекции, нейтрализует токсины. Он имеет две формы: сывороточную и секреторную. Роль IgD достаточно не установлена Возможно, он служит рецептором В-лимфоцитов IgE участвует в связывании антигенов на слизистых оболочках.
Синтез иммуноглобулинов начинается в онтогенезе в разные сроки. Так, синтез IgM обнаруживается на 10-й, а синтез IgG — c 11—12-й недели внутриутробного развития, но уровень его у плода ниже, чем у взрослых организмов.
Сельскохозяйственные животные имеют различия в наборе иммуноглобулинов. У крупного рогатого скота выявлен igG, подразделяемый на два подкласса: IgG1 и IgG2 с периодом полураспада 20—25 дней, — а также IgA и IgM. У овец и коз зарегистрированы три класса: IgG1, IgG2, IgG3; IgA; IgM. У свиней определены три класса: IgG; IgA; IgM.
У лошадей выявлены IgG, IgG(t), IgM, IgA(t),
Использование генетической обусловленности естественной резистентности в практике животноводства. Естественная резистентность, отражающая врожденный иммунитет, является прежде всего следствием видовой невосприимчивости животных, которая формируется в процессе эволюции. Кроме этого, практика животноводства выявила породную и индивидуальную устойчивость некоторых домашних популяций или групп животных в отношении различных инфекций. При этом животные не заболевают даже в очаге массовой инфекции или переносят заболевание в легкой форме.
В основе видовых, породных и индивидуальных свойств естественной резистентности у животных лежит их наследственная обусловленность, которая отражает генетические особенности организма. Повышение уровня естественной резистентности сельскохозяйственных животных может быть следствием целенаправленного отбора и подбора, способствующего распространению и закреплению в популяции желательных генотипов.
Способность животных проявлять повышенную резистентность становится важным селекционным признаком. Известно, что организация промышленных животноводческих комплексов сопровождается увеличением концентрации поголовья на сравнительно малой территории. При этом может создаваться благоприятная ситуация для быстрого распространения инфекций (или инвазий). В условиях современной промышленной технологии животноводства повышенная резистентность животных приобретает особенно важное значение.
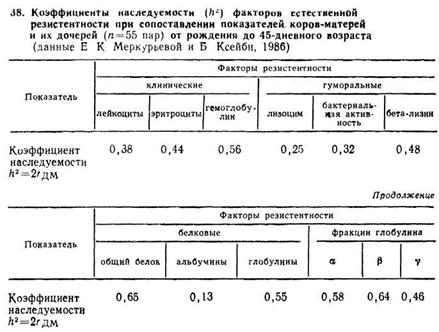
Сила и реакция иммунного ответа на антиген у разных особей данного вида различны и обусловлены индивидуальной наследственностью Передача особенностей иммунитета от родителей потомкам наследуется по законам Менделя и может быть охарактеризована популяционными параметрами, такими как коэффициенты корреляции, регрессии, наследуемости, если сравнить показатели факторов иммунитета (естественной резистентности) между родственными животными. В этом направлении было проведено изучение наследуемости 12 показателей, характеризующих естественную резистентность у группы коров-матерей черно-пестрой породы и их дочерей в молозивно-молочный период онтогенеза (Меркурьева и сотр., 1986). Данные, полученные в процессе исследований, приведены в табл. 38.
Из представленных в табл. 38 данных о величине коэффициента наследуемости можно сделать вывод о том, что при проведении селекции в молочном стаде путем отбора более резистентных коров-матерей можно получать более устойчивое к инфекциям потомство.
Для формирования иммунологического гомеостаза желателен такой тип нервной деятельности и поведения животных, который мог бы поддерживать требуемый гомеостаз и иммунную реактивность, обеспечивающую защиту организма от инфекционного воздействия и влияния неблагоприятных условий среды. Поэтому осуществление селекционой работы по созданию желательного типа высшей нервной деятельности и поведения способствует также и закреплению в породе, линиях или семействах естественной резистентности и иммунной защищенности.
Генетически обусловленная резистентность к болезням и устойчивость к неблагоприятным условиям среды должны стать элементом оценки животных и отражаться в планах племенной работы со стадом и породой. Разработка методов оценки генетической обусловленности резисгентности животных является важным разделом современной зоотехнии
Классификация наследственных патологических отклонений. Аномалии или болезни животных вызываются эндогенными (наследственность) и экзогенными (условия окружающей среды) факторами. С учетом этих факторов выделяют три группы болезней и аномалий.
Первая группа болезней и аномалий вызывается мутацией генов одного или нескольких локусов с наследованием в виде простых менделевских закономерностей (аномалии, уродства, гемофилия и т. д ).
Вторая группа болезней обусловлена взаимодействием наследственности с факторами среды. Наследование их носит полигенный характер и находится под влиянием генов-модифика-горов. При этом признак характеризуется специфической прерывистостью его фенотипического проявления, получившей название порогового состояния признака, например резистентные — восприимчивые, выживающие — погибающие, больные — здоровые. Такое фенотипическое пороговое состояние признака наблюдается при определенном числе активных генов и их кумулятивном действии и имеет разную силу выраженности (экспрессивности) или проявляется в виде пенетрантности, когда заболевание охватывает часть членов популяции.
Болезни третьей группы обусловлены воздействием неблагоприятных факторов среды. Такие заболевания протекают на фоне модификационной (ненаследственной) изменчивости, но при этом реакция разных особей на изменение условий будет неодинаковой, что зависит от генотипа конкретного организма. У животных известен ряд уродств, вызываемых условиями среды; их называют фенокопиями, так как фенотипически эта группа уродств или аномалий сходна с теми, что вызывают мутации, изменяющие наследственность, но не передающиеся потомству. Например, в птицеводстве при нарушении режима инкубации яиц наблюдаются уродства цыплят, подобные наследственным.
Методы определения наследственной обусловленности аномалий и болезней. Для определения наследственной обусловленности зарегистрированной аномалии или заболевания используют комплекс зоотехнических, генетических и ветеринарных методов.
Зоотехнический метод основывается на анализе родословной животного, у которого обнаружено уродство или заболевание. Для этого в группе предков животного, братьев, сестер и боковых родственников устанавливают, была ли у кого-либо из них аналогичная патология или нет; выявляют связь обнаруженной патологии с определенным предком, послужившим родоначальником патологического эффекта; проводят оценку производителей по фенотипу, родословной и по качеству потомства.
Генетические методы включают специальный подбор пар. на основе которого осуществляют анализирующее скрещивание и семейный анализ. К генетическим методам относят цитогенетическую характеристику кариотипа с целью выявления хромосомных аномалий. Применяют иммуногенетические методы, позволяющие оценить иммунную совместимость или ее отсутствие у родителей. Генетико-стагистический анализ популяции дает возможность установить степень гомо - и гетерозиготности локусов по аномальным генам, определить частоту летального аллеля и сделать прогноз вероятности его распространения.
Ветеринарные методы используют показатели клеточного и гуморального иммунитета, патологоанатомический анализ для суждения о патологии и аномалиях у конкретной особи или в обследуемой группе животных. Комплексный подход при выявлении наследственной обусловленности и типа наследования различных аномалий и болезней ставит задачу генетической диагностики, профилактики и разработку методов лечения или ослабления патогенетического эффекта наследственных болезней.
Основные типы аномалий и наследственных заболеваний. У сельскохозяйственных животных выявлено более 130 наследственных аномалий и заболеваний, имеющих генетическое происхождение. Большая часть их затрагивает морфологическое строение, выражаясь в аномалиях скелета, кожи, головного мозга, органов зрения, пищеварении, мышечной ткани, половой и мочевыделительной систем, синтеза пигмента, в аномалии обмена веществ. К таким аномалиям и болезням относятся: водянка головного мозга, аномалии скелета, крипторхизм, гермафродитизм, катаракты, альбинизм, аномалия зубной системы, дисплазия центральной нервной системы (тремор, атаксия, эпилепсия, параличи), карликовость, дисплазия коленной чашечки и тазобедренного сустава, мышечная дистрофия, грыжа (пупочная, паховая, мошоночная) аномалии кровеносной системы и крови (гемофилия) заболевания щитовидной железы (зоб, микседема, гипертиреоз), диабет Созданы международная классификация и список летальных дефектов (по Стормонту и Визнеру) У крупного рогатого скота учтено 46 аномалий и заболеваний, у лошадей 10, у свиней 18, у овец 15, у кур 45, у индеек — шесть, у уток три, у голубей — три.
Полученные в различных опытах данные свидетельствуют о сходном действии ряда генов в организме животных разных видов, вызывающем одинаковые аномалии и болезни. Рецессивный характер наследования приводит к летальному или полулетальному исходу в эмбриональный или постэмбриональный периоды развития особи.
Селекция на устранение из популяции наследственных аномалий и дефектов менее сложна, чем на повышение естественной резистентности, так как фенотипическое проявление аномалий или уродств выявляется при гомозиготном состоянии рецессивного гена, обусловливающего патологию. Такую патологию легко обнаружить в стаде по фенотипическому проявлению аномалий, которые отмечают в редких случаях. Для предотвращения дальнейшего распространения аномалий в поколениях проводят выбраковку животных, проявляющих уродство, или их родителей, через которых они передаются, в результате чего популяция очищается от носителей генетической патологии.
Исследования по изучению аномалий и уродств проведены А И Жигачевым на десяти племпредприятиях, разводящих молочный скот, в ряде зон и областей РСФСР, с использованием фенотипического описания животных, кариотипического исследования, анализа родословных и популяционного анализа (1986). Установлено, что средняя частота встречаемости аномалии невысокая (1,15% — в костромской породе), но у сычевского и симментальского скота встречаемость транслокаций достигает 8,2 — 26,66%.
Анализ спектра аномалий и динамики в их частоте указывает на наличие генетического груза, установленного в процессе мониторинга по поколениям и по календарным отрезкам. Исследование показало необходимость цитогенетической аттестации животных и проведения семейного и общепопуляционного анализа в породах и стадах для планирования мер, пресекающих распространение аномалий.
У крупного рогатого скота выявлены следующие аномалии: доминантные — ахондроплазия (бульдоговидные телята); рецессивные — бесшерстность телят (летальный исход), отсутствие конечностей, укорочение позвоночника (мертворождение), общая водянка, анкилоз суставов, смещение зубов (летальный исход), атрезия (отсутствие) ануса, мозговая грыжа, укорочение или отсутствие нижней челюсти, гидроцефалия, врожденные судороги (летальный исход), удлинение срока стельности на 20 — 90 дней (мертворождение) и на 80 — 100 дней (извлечение плода хирургическим путем), дисфункция щитовидной железы (гибель через две недели после рождения), гиперемия кожи и слизистых оболочек, выкидыши.
У крупного рогатого скота сцепленные с полом доминантные признаки, имеющие летальный характер, приводят к гибели бычков, отсутствию у них зубов, волосяного покрова, к недоразвитию передней доли гипофиза. Действие доминантных генов при их неполной пенетрантности сопровождается аномалией черепа и гибелью животного.
У свиней выявлены рецессивные (мозговая грыжа, отсутствие ануса, недоразвитие ушных раковин, уродство или паралич конечностей, водянка мозга, микседема, выпадение прямой кишки) и доминантные аномалии (порфирия — красно-коричневая окраска костей и зубов, гемофилия, желтуха новорожденных).
У овец зарегистрированы аномалии, обусловленные рецессивным действием генов: мышечная контрактура и мертворожденность, недоразвитость ушной раковины, паралич задних конечностей, деформация скелета, грыжи, отсутствие фаланг, летальная серая окраска шерсти у каракульских овец, карликовость, патологическая светочувствительность, мышечная дистрофия, приводящая к гибели вскоре после рождения, отсутствие нижней челюсти и непроходимость пищевода, отсутствие ануса.
У лошадей рецессивные аномалии выражаются в виде непроходимости ободочной кишки; дефектов эпителия кожи; искривления грудных конечностей; мозжечковой атаксии (опрокидывание на спину, паралич и гибель на 5—6-й день); отсутствия глазного яблока, грудных конечностей; пупочной грыжи; искривления шеи.
Подробный материал по аномалиям и болезням собак приведен в книге R. Robinson (1982), в которой дается их генетический анализ. Даны перечень и характеристика 57 заболеваний и аномалий, в том числе 15 — с доминантным проявлением. Выявлено большое разнообразие заболеваний, затрагивающих различные системы жизненно важных органов и систем. К числу доминантных патологий относятся катаракта, дисплазия бедра, гемофилия, бесшерстность, лимфоотек, микрофтальмия.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
