
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Пример. Необходимо определить среднюю скорость молокоотдачи у коровы, если за 3 мин выдоено 6 кг молока, в том числе за первую минуту — 2 яг, за вторую — 3 кг, за третью — 1 кг
Используем указанную формулу
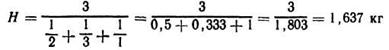
Если же вычислить простую среднюю арифметическую, то скорость молокоотдачи составит:
![]()
то есть полученная величина недостоверна.
Мода и медиана. Дополнительными характеристиками среднего значения варьирующего признака служат мода (Мо) и медиана (Me). Мода показывает, какая величина варианта (хМо) данного признака чаще всего встречается в совокупности. Медиана указывает на то, какой вариант расположен в середине (центре) вариационного ряда; этот вариант делит совокупность на две равные части: с уменьшающимися и увеличивающимися значениями х от медианы. Использование моды и медианы особенно удобно для сопоставления совокупностей по качественным признакам. Например, модальной мастью для скота холмогорской породы будет черно-пестрая; модальное число сосков у коров — четыре, у свиней — 10 и т. д.
Между шестью рассмотренными параметрами средних величин существует определенное соотношение» а именно:
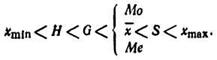
Следовательно, величина средней квадратической S всегда больше, а величина средней гармонической H всегда меньше любой другой средней. В нормальном распределении величины ![]() , Mo, Me совпадают. Неправильно выбранный параметр искажает истинную среднюю величину признака.
, Mo, Me совпадают. Неправильно выбранный параметр искажает истинную среднюю величину признака.
Вычисление степени изменчивости признаков. Наиболее простой способ установления изменчивости признака в совокупности — определение минимальной (xmin) и максимальной (xmax) величин вариантов, то есть определение лимита. Чем больше абсолютная разность между xmax и xmin, тем более значительна изменчивость признака.
Среднее квадратическое отклонение. Основным показателем изменчивости служит величина а. Формула ее имеет следующий вид:
![]()
Наличие в формуле квадратного корня со знаками «+» и «—» указывает на то, что величина этого параметра характеризует изменчивость признака особей совокупности относительно средней арифметической, то есть варьирование в любой совокупности выражается отклонением вариант в сторону «+» или «—» от ![]() . Под корнем в знаменателе ставят n или число степеней свободы (v=n - 1), что дает так называемую несмещенную величину σ.
. Под корнем в знаменателе ставят n или число степеней свободы (v=n - 1), что дает так называемую несмещенную величину σ.
Среднее квадратическое отклонение не только характеризует изменчивость, но и выявляет особенности варьирования признака у особей совокупности. Например, может быть такая ситуация, когда две сопоставляемые совокупности имеют одинаковые величины xmax, xmin и ![]() , но по особенностям варьирование и величине σ различаются.
, но по особенностям варьирование и величине σ различаются.
Варьирование признака членов совокупности может быть нормальным, асимметричным, эксцессивным, биномиальным, пуассоновым, трансгрессивным. Наиболее распространено нормальное варьирование.
Установлено, что в выборках, которые характеризуются нормальным распределением, весь размах изменчивости ограничивается значениями от xmin= —Зσ до xmax = +3σ от средней арифметической, то есть весь лимит включает 6σ. В границах х±3σ сосредоточено 99,7% всех членов совокупности и только 0,3% членов совокупности выходят по величине признака за пределы —Зσ и +3σ. Закономерности нормального распределения выражаются тремя функциями. Так, если σ удоев равна 500 кг, а ![]() = 3000 кг, то минимальный удой (xmin) у корой в такой совокупности, вероятнее всего, будет равен
= 3000 кг, то минимальный удой (xmin) у корой в такой совокупности, вероятнее всего, будет равен ![]() —3σ=3000—3·500=1500 кг, а максимальный (xmax) = х+3 σ = 3000+3·500=4500 кг.
—3σ=3000—3·500=1500 кг, а максимальный (xmax) = х+3 σ = 3000+3·500=4500 кг.
Коэффициент вариации. Кроме лимита и а, являющихся основными показателями изменчивости, используют и Коэффициент вариации (Сv), который выражает степень изменчивости признака в процентах от величины средней арифметической. Формула коэффициента вариаций следующая:
![]()
Еслн число членов выборочной совокупности более 30 (п>30), а возможность применения счетно-вычислительной Техники ограничена, то дли определения средних величин (![]() ), показателей изменчивости (σ и Сv }) применяют метод упорядочения данных. Для этого составляют так называемые вариационны ряды, где величину варьирующего признака (хi) оформляют в виде классов с переходом от меньшей к большей величине признака, а членов совокупности распределяют по классам и выявляют частоты рi (табл. 5).
), показателей изменчивости (σ и Сv }) применяют метод упорядочения данных. Для этого составляют так называемые вариационны ряды, где величину варьирующего признака (хi) оформляют в виде классов с переходом от меньшей к большей величине признака, а членов совокупности распределяют по классам и выявляют частоты рi (табл. 5).
Обработку вариационных рядов осуществляют разными способами. В данном примере Использован метод произведений. Для вычисления средней арифметической умножают величину класса (х) на соответствующую ему величину частоты (p), при этом получают ряд значений ![]() ;
;![]() позволяет определить среднюю арифметическую плодовитость лисиц;
позволяет определить среднюю арифметическую плодовитость лисиц;
![]()
Для вычисления σ выражают порядковым числом (1, 2, 3...) отклонение каждого класса oт класса, взятого за нулевой (класс условной средней Ах). В качестве нулевого принят класс с наибольшей величиной p, расположенный в центре ряда. В табл. 5 этому условию соответствует величина x0 = 5 голов.
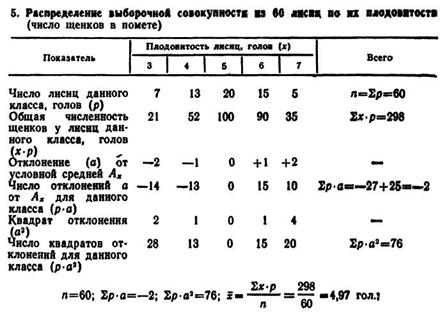
От этого класса в сторону, как увеличения, так и уменьшения признака записывают условное порядковое отклонение (—2; —1; 0; +1; +2). В четвертой строке табл. 5 записывают число отклонений для данного класса (р·а), в пятой строке — квадрат отклонения (а2). В шестой строке записывают результаты умножения (р·а2). Полученные данные позволяют вычислить, а по следующей формуле:
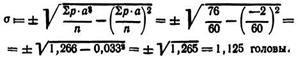
Коэффициент вариации вычисляют по формуле
Сv=(σ: ![]() ) X100= (1,125: 4,9=22,6%.
) X100= (1,125: 4,9=22,6%.
Следовательно, в данном примере среднее квадратическое отклонение по плодовитости лисиц составит 1,125 головы при степени изменчивости признака 22,6%.
Варианса. Для анализа изменчивости признака важное значение имеет так называемая варианса, которую получают возведением среднего квадратического отклонения в квадрат. Показатель вариансы (σ2) используют в генетическом анализе, когда требуется разложить фенотипическую изменчивость на составляющие ее части: изменчивость, обусловленную разнообразием генотипов особей совокупности, и изменчивость, обусловленную влиянием различных факторов среды. Соотношение варианс фенотипической (σф2), генотипической (σг2) и паратипической (σn2) можно записать так: σф2= σг2 + σn2.
Нормированное отклонение. В изложенном выше речь шла о характеристике всех членов совокупности по показателю изменчивости признака, но можно выявить и особенности различных признаков отдельно взятого члена совокупности, используя в качестве критерия нормированное отклонение (t). Для этого величину признака данной особи определяют как отклонение от средней арифметической, а полученное отклонение (xi— ![]() ) делят на величину а, то есть t= (xi —х) : σ. В результате получают относительную оценку признака в долях σ. Данным способом можно сопоставлять разные признаки одной или разных особей.
) делят на величину а, то есть t= (xi —х) : σ. В результате получают относительную оценку признака в долях σ. Данным способом можно сопоставлять разные признаки одной или разных особей.
Например, среднегодовая продуктивность в стаде — 5500 кг, средняя жирность молока — 3,6%, средние квадратические отклонения удоя и жирномолочности (σy и σх) соответственно 600 кг и 0,1%. Для сопоставления отобраны две коровы с рекордной продуктивностью: № 1—6500 кг; 3,8% жира и № 2—7000 кг; 3,5% жира. Следовательно, в первом случае:

Сопоставление животных показало, что они заметно различались по отклонению от средних по продуктивности, особенно по содержанию жира в молоке.
Определение величины и направления связей между признаками. Основным биометрическим показателем, позволяющим определять величину и направление связи между признаками, служит коэффициент корреляции (r). Он показывает величину связи между двумя, тремя и большим числом признаков. Величина этого коэффициента принимает дробное выражение в пределах от 0 до ±1. Чем ближе показатель к единице, тем больше связь между коррелирующими признаками. Наличие знака «+» означает, что между признаками существует положительная корреляция, когда при увеличении одного признака у особей совокупности другой признак также возрастает или при уменьшении одного признака другой также снижается. Следовательно, изменение признаков происходит в одном направлении. Если же коэффициент корреляции имеет знак «—», то это указывает на наличие между признаками отрицательной (обратной), разнонаправленной связи, при которой увеличение одного признака сопровождается уменьшением другого.
Упорядочение первичных данных для вычисления r осуществляют с помощью корреляционной решетки (рис. 53), в которой верхнюю строчку и левую боковую графу заполняют классами коррелируемых признаков (х и у). В клетки, образуемые пересечением граф и строк, разносят данные о членах совокупности с учетом обоих признаков; в результате выявляются частоты (р) в каждой клетке. В зависимости от степени и направления корреляции распределение частот в клетках корреляционной решетки может быть различным.
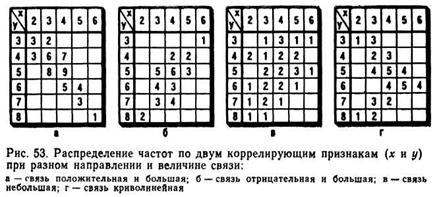
Если корреляция между признаками положительная и большая, то наблюдается накопление частот по диагонали из верхнего левого к нижнему правому углу решетки (см. рис. 53, а). При отрицательной корреляции частоты распределяются по противоположной диагонали (см. рис. 53, б). Если корреляция между признаками незначительна, то частоты распределяются по большей части клеток (см. рис. 53, в). В тех случаях, когда корреляция между признаками криволинейна, частоты распределяются так, как показано на рис. 53, г. При криволинейном типе корреляции один из признаков увеличивается, а другой, связанный с ним, сначала возрастает (или уменьшается), затем снижается (или повышается).
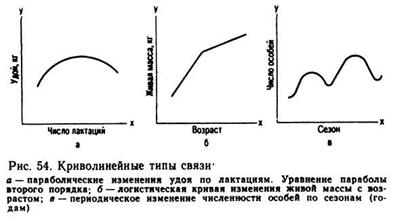
При криволинейной связи коэффициент корреляции не может отразить ее наличие; он или уменьшает ее величину, или вовсе не обнаруживает. Например, если графически выразить зависимость между возрастом коров и их удоями за лактацию, то изменение удоя выразится кривой (рис. 54), которая показывает, что до определенного возраста удои увеличиваются, а затем уменьшаются. В первой части кривой связь между удоем и возрастом большая и положительная, во второй части кривой связь отрицательная. Следовательно, коэффициент корреляции будет близок к нулю, связь как бы отсутствует, а в действительности она имеется (и даже значительная), но не улавливается этим статистическим коэффициентом, так как +r и —r «гасят» друг друга.
Связь признака с возрастом обычно имеет криволинейный характер, и поэтому пользоваться коэффициентом корреляции в этих случаях нецелесообразно, так как r будет близок к нулю или приуменьшит истинную связь между возрастом и варьирующим признаком.
В основе формул, позволяющих вычислить величину коэффициента корреляции, лежат следующие показатели признаков х и  С помощью этих величин можно определить r для малых выборок, а при использовании счетно-вычислительных машин — и для больших выборок. Техника вычислений сводится к составлению рядов из величин x; y; х·у; х2 ;у2 и их суммирования по графам.
С помощью этих величин можно определить r для малых выборок, а при использовании счетно-вычислительных машин — и для больших выборок. Техника вычислений сводится к составлению рядов из величин x; y; х·у; х2 ;у2 и их суммирования по графам.
Рабочие формулы для определения r могут быть различными, поэтому выбирают наиболее удобные с учетом абсолютной величины коррелируемых признаков. В основную формулу коэффициента корреляции введены показатели нормированного от клонения признака каждого члена совокупности от средней арифметической:
![]()
В данном случае σх и σу вычисляют без умножения на классовый промежуток.
Для малых выборок удобнее пользоваться формулой
Если требуется определить коэффициент корреляции на большой выборке с применением корреляционной решетки, то пользуются следующей формулой:
![]()
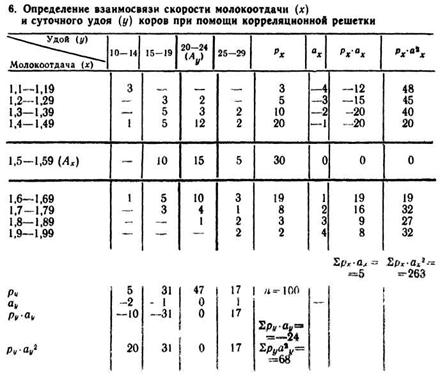
где р — частоты по клеткам решетки, σх и σу средние квадрат ические отклонения для каждого коррелируемого признака, выраженные в относительных, а не в именованных величинах; п — объем выборки, величины bх и by — поправки для каждого ряда
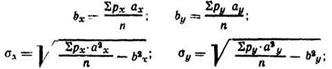
ах и ау — условные отклонения от нулевого класса, у которого отмечено наибольшее число частот. Этот класс расположен близко или совпадает с классом, занимающим центральное положение.
В качестве примера рассмотрим определение корреляции между скоростью молокоотдачн (кг/мин) и суточным удоем у коров черно-пестрой породы. Обследовано 100 коров, данные о которых распределили следующим образом (табл. 6).
По горизонтали расположен вариационный ряд суточного удоя (у), по вертикали — ряд скорости молокоотдачи (x). Выделены классы условной средней Ау=20—24 кг и Ax = 1,5—1,59 кг/мин, заполнен ряд условных отклонений ах и аy. На эти величины умножают соответствующие частоты ряда и получают значения рх-ах и py-ay. Затем, умножая рхах на отклонение ах, получают ряд рх-а2х; соответственно ![]() Из этого следует:
Из этого следует:
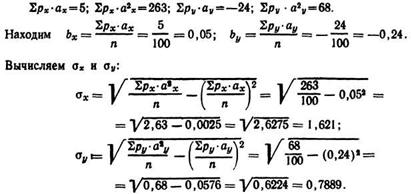
Затем определяют величину![]() Для этого в углах (квадрантах),
Для этого в углах (квадрантах),
отграниченных «крестом» нулевых классов, умножают частоту (р) каждой клетки на соответствующее значение ах и ау:

При вычислении I и IV квадранты всегда дают положительные, а II и III — отрицательные величины ![]() Подставив полученные
Подставив полученные
величины в формулу, определяют r:
![]()
Следовательно, связь между удоем и скоростью молокоотдачн положительная.
Коэффициент регрессии (b) показывает, насколько изменяется признак у, если х изменяется на определенную величину. Коэффициент регрессии — величина именованная. Он может иметь два значения, то есть показывать изменение признака х по у и, наоборот, у по х. Коэффициент регрессии при прямолинейном типе связи рассчитывают с применением величин r;
![]()
Регрессия между признаками может быть выражена в виде эмпирического и теоретического рядов регрессии, в виде графика, а также через уравнения регрессии.
Например, изменение скорости молокоотдачи (х) при изменении суточного удоя (у) составляет эмпирический ряд регрессии х по у, который выразится следующим образом (см. табл. 6). Для получения эмпирического ряда х по у умножают частоты каждой клетки решетки по классу у на середину класса по х, суммируют эти данные и делят на все частоты класса
у, то есть ![]() . Так, для класса с удоем 10—14 кг получают следующее среднее значение скорости молокоотдачн:
. Так, для класса с удоем 10—14 кг получают следующее среднее значение скорости молокоотдачн:
![]()
Аналогичный расчет делают для остальных трех классов:

Регрессию скорости молокоотдачи можно представить таким образом:

Уравнение регрессии при прямолинейном типе связи выражается формулой x=bу+a или у=bх+а. Изменение х по у определяется уравнением х=bу+а, где х — искомая зависимая функция (зависимый признак); у — аргумент (независимый признак); b — коэффициент связи между х и у; а — исходный уровень зависимого признака, если y=0.
Уравнение может иметь следующий вид;
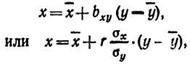
где ![]() —средняя арифметическая зависимого признака, соответствующая значению a; bxv соответствует значению
—средняя арифметическая зависимого признака, соответствующая значению a; bxv соответствует значению![]() в уравнении.
в уравнении.
Если зависимым признаком будет у, а независимым х, то в уравнении произойдет замена мест между значениями признаков:
![]()
Пользуясь уравнениями регрессий, можно составить теоретический ряд значений и построить график для х при изменении у или у по х (рис. 55).
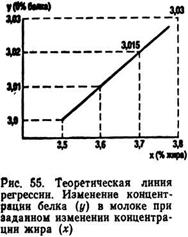
Коэффициент корреляции между альтернативными признаками rа (не только количественными, но и качественными) особенно удобен при анализе генетических данных. Для его определения первичные данные по каждому члену выборки размещают в четырехклеточной корреляционной решетке. Формула для определения ta выглядит так:
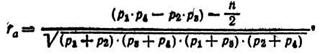
где p1, p2, p3, p4 — частоты, распределившиеся в четырех клетках альтернативных признаков корреляционной решетки.
Величина rа может находиться в пределах от —1 до +1.
Например, при проведении экспериментального скрещивания кур нескольких пород требовалось определить по соотношению окраски пуха у цыплят, происходило ли сцепленное с полом наследование «сигнальной» окраски.

В опыте было получено 100 помесных цыплят, которые распределились в клетках корреляционной решетки следующим образом (табл. 7). Величину rа вычисляют по формуле
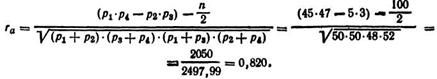
Следовательно, связь окраски пуха цыплят с половой принадлежностью очень высокая, что позволяет получить в опыте аутосексную группу птицы.
Полихорический коэффициент связи. В практике селекционной работы и при анализе генетических данных иногда необходимо установить связь между качественными признаками, которые оценивают «на глаз», грубо, но зоотехнически вполне допустимо. Например, связь между конституцией животных и степенью упитанности, конституцией и формой завитка смушка овец или связь признаков потомства с признаками родителей и т. п. В таких случаях связь устанавливают с помощью полихорического коэффициента связи ρ, который высчитывают по формуле
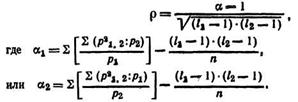
Здесь i1 и i2 — число классов по каждому признаку; p1,2 - частоты в клетках корреляционной решетки; р1 и р2 — частоты вариационного ряда каждого из признаков; n — число членов в выборке.
Формула удобна тем, что при использовании ее нет необходимости вычислять σ1 и σ2, что значительно упрощает расчеты. Величина ρ колеблется от 0 до 1, но этот коэффициент не выявляет направления корреляции, а только указывает на ее величину, поэтому он всегда имеет положительный знак. Полихорический коэффициент применяется и в тех случаях, когда один признак имеет несколько градаций.
Пример. Требуется определить величину связи между типом конституции коров-матерей и их дочерей. Обследовано 50 пар (мать — дочь). Пары распределены по клеткам корреляционной решетки (табл. 8).

Каждую частоту в клетке решетки обозначают p1.2, возводят это число в квадрат (p1.2)2 и затем делят на значение частот, размещенных в строчке р2. Проведенная обработка дает для каждой клетки выражение (р1.2)2: p2.
Полученную величину суммируют, получая![]() и записывают сумму в
и записывают сумму в
графу, идущую после графы с частотами вариационного ряда р1. Проведя вычисления по формуле![]() получают итоговое число 2,23. Подставляют данные в формулу a1:
получают итоговое число 2,23. Подставляют данные в формулу a1:
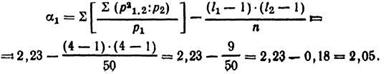
Используя величину a1, находят полихорический коэффициент
![]()
Для данного примера на основании величины ρ можно сделать вывод, что тип конституции дочерей коррелирует с типом конституции матерей, а полихорический коэффициент равен 0,35, то есть наследственно обусловлен и может быть выражен коэффициентом наследуемости h2—2r, В нашем примере h2 =2 ·0,35 = 0,70 выявляет наследственную обусловленность конституции потомства.
Ранговый коэффициент связи по Спирмену (rs). При обработке первичных материалов могут встретиться признаки, которые нельзя измерить ни точно, ни грубо, поэтому их выражают порядковым местом (рангом). Например, определяют ранга быков-производителей по удою (х) их дочерей и сопоставляют данные с рангами их дочерей по показателю живой массы (у). Ранговый коэффициент корреляции выражают следующей формулой:
![]()
где n —число сопоставляемых пар рангов; D — разность между парами рангов признака х с рангами признака у, Величина ra изменяется от —1 до
Пример. Необходимо определить, имеется ли связь между ростом (х) рысистых лошадей и скоростью их бега (у) на дистанцию 1600 м. Сравнивают ранги пяти лошадей по этим двум признакам.
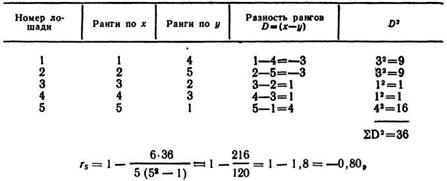
то есть связь между ростом и скоростью бега лошади на дистанцию большая и обратная: у лошадей более высокого роста в среднем скорость бега выше, чем у лошадей низкого роста.
Криволинейные связи, корреляционное соотношение. Кроме прямолинейных или близких к ним связей, для определения которых можно применять перечисленные коэффициенты, существует многообразие связей криволинейного типа. С целью их установления используют другие формулы и коэффициенты. Многие признаки сельскохозяйственных животных проявляют криволинейную связь с возрастом. Изменение живой массы животного от рождения до наступления половой зрелости выражается так называемой логистической кривой. У насекомых наблюдаются периодические вспышки усиления и затухания в размножении, что можно выразить периодической синусоидной кривой.
При наличии криволинейного типа связи между признаками, как правило, используют показатель корреляционного соотношения ![]() который позволяет определить величину связи, но не выявляет направления ее, то есть этот коэффициент всегда положительный. Его величина изменяется от 0 до + 1. Корреляционное соотношение можно применять для измерения как криволинейных, так и прямолинейных связей.
который позволяет определить величину связи, но не выявляет направления ее, то есть этот коэффициент всегда положительный. Его величина изменяется от 0 до + 1. Корреляционное соотношение можно применять для измерения как криволинейных, так и прямолинейных связей.
При криволинейном типе связи величина ![]() всегда будет больше величины r, вычисленной на том же материале. Если же связь прямолинейна, то
всегда будет больше величины r, вычисленной на том же материале. Если же связь прямолинейна, то ![]() = r. Корреляционное соотношение имеет два значения: у по х и х по у, следовательно, оно выявляет неравенство связей, так как
= r. Корреляционное соотношение имеет два значения: у по х и х по у, следовательно, оно выявляет неравенство связей, так как
В общем, виде формула корреляционного соотношения включает показатели групповых (частных) средних квадратических и общих средних квадратических отклонений:
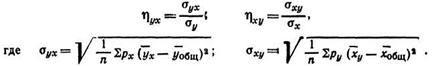
Для вычисления корреляционного соотношения по вариационным рядам, оформленным с распределением признаков на варьирующие классы (то есть при взвешенных рядах), пользуются следующей формулой:
![]()
где ![]() — среднее арифметическое признака у;—
— среднее арифметическое признака у;— ![]() средние величины
средние величины
признака у по классам признака х. Здесь зависимой функцией является признак у, а независимым аргументом — признак х,
Для определения ![]() ух составляют корреляционную решетку и определяют эмпирические величины регрессии признака у в зависимости от признака х.
ух составляют корреляционную решетку и определяют эмпирические величины регрессии признака у в зависимости от признака х.
Пример. Определение влияния возраста (х) свиноматок на их плодовитость (у). Возраст выражен числом опоросов. В выборку вошло 75 опоросов (п), полученных от 50 свиноматок, и учтено для каждого опороса число родившихся поросят. Вначале необходимо составить корреляционную таблицу зависимости у от х (табл. 9).

Вычисляют эмпирические величины регрессии изменения признака плодовитости (у) от изменения возраста (х). Для этого умножают частоты клеток решетки (рух) на соответствующую величину признака у каждого класса, то есть по столбцам, и для каждого столбца записывают получаемую сумму от умножения рух-у и делят на число pxi:
1-й столбец: (6·8+4·11) : (6+4+44) : 10=92: 10=9,2;
2-й столбец: (3·8+6·11 + 1·14) : 10=10,4 и т. д.
Полученный ряд изменений у по х демонстрирует эмпирический ряд регрессии, из которого видно, что с 3-го опороса по 6-й наблюдается повышение плодовитости свиноматок, а с 7-го — снижение.
Определяют среднюю плодовитость свиноматок по всем возрастным группам, то есть уобщ. Для этого перемножают частоты ряда у(ру) по каждому классу на середину класса. Суммируют данные по всем четырем классам признака плодовитости и, деля сумму на общее число обследованных опоросов (n=75), получают среднюю плодовитость по всему корреляционному комплексу. Эти данные введены в табл. 10. Средняя плодовитость
![]()
Далее обработка данных сопровождается получением суммы квадратов отклонения для показателей y от общей средней и умножения на частоты ряда у, то есть![]() и вычисление суммы квадратов отклонений
и вычисление суммы квадратов отклонений
частных средних ![]() от общей средней, умноженных на частоты для
от общей средней, умноженных на частоты для
эмпирического ряда регрессии, то есть на рх, в соответствии с выражением  Эти промежуточные расчеты приведены в табл. 11 и 12.
Эти промежуточные расчеты приведены в табл. 11 и 12.
Далее определяют ![]() где
где ![]() — средняя плодовитость по столбцам, получена в показателях эмпирической регрессии
— средняя плодовитость по столбцам, получена в показателях эмпирической регрессии![]() 9,2; 10,4; 12,4; 12,8; 12,2; 13,0; 11,0; 11,0).
9,2; 10,4; 12,4; 12,8; 12,2; 13,0; 11,0; 11,0).
Полученные величины вводят в формулу корреляционного соотношения при обработке взвешенных рядов:
![]()
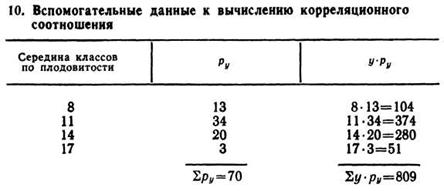

Полученная величина ηух указывает на большую зависимость плодовитости от возрастного показателя свиноматок, что следует из ряда регрессии у по х.
Генетический коэффициент корреляции между признаками. В практике селекции немаловажное значение имеет определение связей между признаками, обусловленными наследственностью. В основе выявления генетической детерминации наследования признаков лежит генетический анализ, при котором признаки потомков сравнивают с признаками их родителей. Такой анализ можно осуществлять в отношении качественных признаков, которые легко прослеживаются у родителей и потомков данной семьи.
Если же необходимо определить наследование количественных признаков и их генетическую обусловленность, то для анализа недостаточно изучения данных в пределах одного семейства, а потребуется его осуществление на массовом материале с применением популяционного и математического методов. Для этого применяют метод определения генетического коэффициента корреляции (rG) между признаками, разработанный Хейзелем (1943). Суть метода заключается в том, что на группах родственных животных (матери — дочери, отцы — сыновья, полусестры) вычисляют четыре коэффициента корреляции между двумя разными фенотипическими признаками (х и у) в пределах каждой сопоставляемой родственной группы и между группами.
Формула Хейзеля для генетического коэффициента связи между признаками х и у построена на определении фенотипических коэффициентов корреляции между х и у, учтенных у животных родственных групп, например у матерей и дочерей. Это означает, что вычисляют фенотипические коэффициенты корреляции между признаками х дочерей и матерей, у дочерей и матерей, х дочерей и у матерей, у дочерей и х матерей. Данные берут из первичного зоотехнического учета. В результате получения четырех величин r определяют генетический коэффициент связи между признаками х н у, используя следующую формулу:
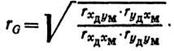
Формулу применяют для тех случаев, когда оба r в числителе имеют знак «+» или знак «—», но в знаменателе оба должны быть положительными. Если же под корнем в числителе один из г имеет знак «—», а другой — знак «+», то формула видоизменяется:
![]()
Наличие отрицательной связи между хд и хм или между yд и уи указывает на сильное взаимодействие генотипа со средой или на сложный тип наследования (эпистаз, межаллельное взаимодействие), и, следовательно, по формуле Хейзеля нельзя выявить связь, так как формула основана на предположении о наличии аддитивного действия генов коррелирующих признаков.
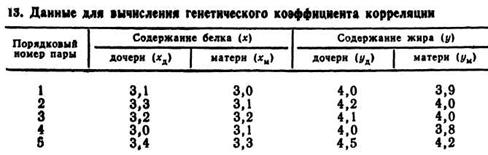
Пример. Необходимо вычислить генетический коэффициент корреляции между содержанием жира и белка в молоке коров-дочерей и их матерей. Исходные данные для пяти пар «мать — дочь» приведены в табл. 13.
Методом малой выборки определяют r xдxм, r yдyм, r xдyм, r yдxм,
Способом, указанным на с. 191, определяют фенотипические коэффициенты корреляции. Из этого следует:
содержание белка (матери — дочери) — r xдxм = 0,65;
содержание жира (дочери — матери) — r yдyм = 0,80;
содержание белка (дочери) — содержание жира (матери)— r xдyм= 0,733;
содержание жира (дочери) — содержание белка (матери) — r yдxм=0,053.
Подставляя эти данные в формулу, получают
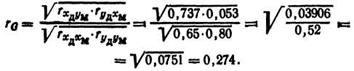
Следовательно, генетическая обусловленность связи между белковомолочностью и жирномолочностью коров выражается rG=0,274, поэтому отбор животных по содержанию жира в молоке будет сопровождаться повышением белковомолочности.
Рассмотренная группа коэффициентов, позволяющих определять величину и направление связи между признаками, показала, что для решения ряда вопросов в зависимости от материала могут быть использованы разные коэффициенты. Их используют для установления сцепленного типа наследования; они находят применение и для осуществления косвенной селекции животных на основе корреляций между различными признаками. Корреляционный и регрессионный анализ имеет важное значение для планирования и прогнозирования уровня того или иного признака.
Типы статистических ошибок. При обработке данных, полученных из зоотехнической документации или из специальных опытов, возможны три типа ошибок: технические (просчеты, описки); ошибки, обусловленные неточностью используемого прибора; статистические. Ошибки, вызванные небрежностью, описками, просчетами, должны быть исправлены; чтобы не допустить их, следует более тщательно обрабатывать данные. Ошибки, появляющиеся вследствие неточности прибора, могут быть также легко устранены. Такие ошибки называют систематаческими. Они не имеют отношения к биометрии.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
