

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
свечение удавалось наблюдать также и в растворах, содержащих окись азота, супероксидные радикалы и какой-нибудь белок. Между тем известно, что при реакции окиси азота и супероксида образуется пероксинитрит, соединение, кстати сказать, очень токсичное;
ON· + ·ОО - → ONOO (пероксинитрит)
По-видимому, именно реакция пероксинитрита с белком приводит к свечению, которое вносит существенный вклад в свечение всего органа.
Надо заметить, что природа процессов, определяющих собственное свечение тканей, может меняться при изменении состояния этой ткани. В опытах того же автора было показано, что у животных с воспалением легких ингибитор NO-синтазы слабо влиял на свечение органа, зато свечение снижалось антиоксидантами - супероксиддисмутазой и ловушками липидных радикалов. Логичен сделанный вывод, что при воспалении на первый план выходят реакции, связанные с активацией клеток-фагоцитов и образованием ими активных форм кислорода, а затем - липидных перекисей, тогда как в норме за свечение ответственны реакции окиси азота. Таким образом, собственное свечение ткани обусловлено реакциями трех типов:
Почему оно "сверхслабое", это свечение клеток и тканей?
Чем же объясняется низкая интенсивность хемилюминесценции, сопровождающей реакции свободных радикалов?
Причин целых три. Во-первых, сама концентрация радикалов в биологических системах очень мала из-за их высокой химической активности, поэтому малы и скорости реакций, сопровождающихся свечением. Во-вторых, не всякое химическое взаимодействие радикалов непременно приводит к образованию электронно-возбужденных молекул продуктов реакции, как это изображено на рис. 3 (4). Напротив, в подавляющем большинстве окислительно-восстановительных взаимодействий между молекулами или радикалами электрон переносится не на уровень возбужденного состояния, я на самый нижний свободный уровень, и последующего высвечивания кванта не происходит. В третьих, даже если и образовалась возбужденная молекула продукта, вероятность того, что высветится квант, а не произойдет растраты энергии в тепло, тоже обычно очень мала.
Две последние причины приводят к тому, что квантовый выход хемилюминесценции в случае, скажем, реакции двух перекисных радикалов составляет всего 10-8-10-10. Это происходит потому, что квантовый выход образования возбужденных молекул продукта

равен всего 10-4-10-5, а квантовый выход люминесценции продукта

составляет для кетонов, образующихся при взаимодействии перекисных радикалов, в свою очередь, тоже около 10-4- 10 - 5.
Вот и выходит, что общий квантовый выход хемилюминесценции составляет всего-навсего 10-8-10-10. Отсюда становится понятной стратегия для использования хемилюминесценции как метода обнаружения радикалов. Для усиления свечения, т. е. QХЛ надо увеличить или QВОЗБ или QЛЮМ или и то и другое. Соединения, которые реагируют с радикалами с образованием возбужденных молекул продуктов, такие как люминол или люцигенин, называют химическими активаторами ХЛ, или хемилюминогенными зондами.
Они как бы увеличивают QВОЗБ. Существуют и такие вещества, которые перехватывают возбужденные состояния продуктов и высвечивают кванты с высоким выходом (т. е. как бы увеличивают QЛЮМ); их можно назвать физическими активаторами ХЛ. Подробнее об активации хемилюминесценции и ее применении мы надеемся рассказать в следующей статье в Соросовском Образовательном Журнале.
Заключение
С развитием техники измерения очень слабых световых потоков стало ясно, что свечение при химических реакциях (хемилюминесценция) - не такая уж экзотика. Слабое свечение сопровождает по существу все химические реакции, идущие с участием свободных радикалов. Собственное свечение животных клеток и тканей обусловлено преимущественно реакциями цепного окисления липидов и реакциями, сопровождающими взаимодействие окиси азота и супероксидного радикала.
В присутствии определенных соединений, обычно называемых в отечественной литературе "активаторами", свечение клеток и тканей может быть усилено на несколько порядков величины. Наибольшее распространение получило измерение хемилюминесценции, связанной с выделением клетками активных форм кислорода (к которым относятся супероксид, гидроксильный радикал, перекись водорода и гипохлорит): хемилюминесценция наблюдается в присутствии активаторов люминола и люцигенина. Активированная хемилюминесценция довольно широко применяется в клиническом биохимическом анализе.
В последнее время все больший интерес привлекает собственное ("сверхслабое") свечение клеток и тканей животных и человека, которое обусловлено реакциями свободных радикалов: радикалов липидов и кислорода, а также окиси азота, - соединениями, играющими огромную роль в жизни организма, а при определенных условиях - и развитии ряда патологических состояний.
Известное с древних времен видимое простым глазом свечение некоторых организмов, например светляка, которое называют биолюминесценцией, также нашло широкое применение в клинических анализах и медико-биологических научных исследованиях.
Метод ДСК
В биологических мембранах липидный слой по всем имеющимся данным представляет собой жидкое тело с вязкостью, близкой к вязкости подсолнечного масла.
Строго говоря текучесть мембраны ограничена внутренней гидрофобной фазой, которая состоит из углеводородных цепей жирных кислот. Эта фаза, однако, не всегда бывает жидкой. При охлаждении до температур ниже 10оС мембраны замерзают, т. е. жидкая фаза затвердевает, приобретая свойства двумерного кристалла.
В мембранах, образованныых синтетическими липидами, фазовый переход из жидкого в твердое состояние может происходить при более высоких температурах, в зависимости от химического состава фосфолипида. В таблице 1 приведены температуры фазовых переходов некоторых синтетических фосфатидилхолинов (лецитинов).
Таблица 1. Температуры плавления некоторых синтетических фосфолипидов
Жирные кислоты | Название остатка жирной кислоты | Сокращённое название фосффолипида | Температура плавления, Tc, oC |
14:0 | Миристоил | ДМЛ | 23 |
16:0 | Пальмитоил | ДПЛ | 41 |
18:0 | Стеароил | ДСЛ | 58 |
18:1 | Олеил | ДОЛ | -21(цис-форма) |
Полное название фосфолипидов: ДМЛ - 1,2-димиристоилфосфатидилхолин (еще одно возможное сокращение - „ДМФХ”) и так далее.
Для изучения фазовых переходов при нагревании используют метод дифференциальной сканирующей микрокалориметрии (сокращенно - ДСК). Не останавливаясь на конструкции прибора, отметим только, что в конечном счете с его помощью записывается кривая теплоемкости, т. е. зависимость теплоемкости липидов или мембран в суспензии от температуры:
| (1) |
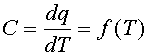
Рис.1. Фазовые переходы в суспензии фосфолипидных везикул (липосом) по данным дифференциальной сканирующей микрокалориметрии (ДСК).
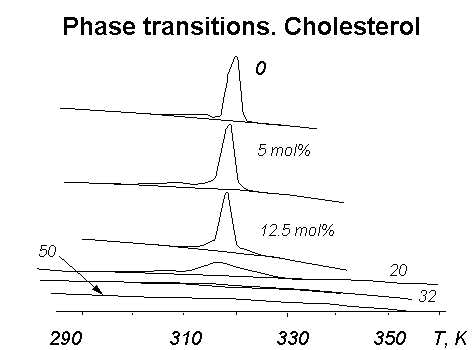
Перед приготовлением липосом к фосфолипидам было дабавлено разное количество холестерина; его содержание в молярных процентах указано у кривых.
По оси ординат отложена теплоемкость, по оси абсцисс - температура, K.
Метод называется дифференциальным, потому что измеряется только теплоемкость суспендированного материала на фоне гораздо большей теплоемкости раствора сравнения.
Примеры таких кривых даны на рис.1, где приведены кривые ДСК для дистеароилфосфатидилхолина (ДСЛ). На этих кривых, в частности, видно увеличение температурного интервала фазового перехода пр добавлении к липиду в мембране холестерина.
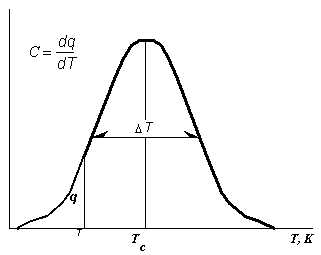
На рисунке 2 слева более подробно обозначены параметры кривой ДСК. На первом этапе нас будут интерессовать три из них:
Температура фазового перехода ("плавления") Tc. Температурный интервал ("ширина") фазового перехода. Общее количество тепла Q, поглощенного при плавлении. Оно представляет собой площадь под кривой ДСК, т. е. функции C=f(T)Рис.2. Характеристик фазовых переходов в липидах по данным дифференциальной сканирующей микрокалориметрии.
С-теплоемкость.
T - полуширина фазового перехода,
Tc - температура плавления.
Кривые плавления
Кривой плавления называется зависимость доли жидкой фазы в общем количестве изучаемого вещества, в данном случае - липидов мембран.
Рис.3. Кривая плавления липиддов в липосомах, приготовленных из ДПЛ.
- доля жидкой фазы, T - температура,
Tc - температура плавления ( = 0,5), ml - количество липида в жидкой фазе, ms - количество липида в твердой фазе.
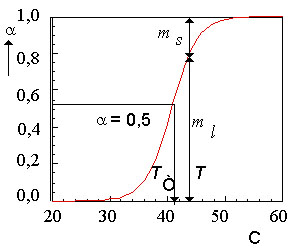
Обозначим количество липидов в жидкой фазе через ml, а количество липидов в твердой фазе через ms. Тогда доля жидких липидов будет равна:
| (2) |
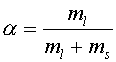
На рис. 3. приведена кривая плавления дистеароил-фосфатидил холина (или дипальмитоил-лецитина, ДПЛ).
Методы измерения кривых плавления
Для определения доли жидкой фазы в общем объеме изучаемого материала, в нашем случае, - в липидной слое мембран, можно использовать разные методы.
Анализ кривых ДСК
Один из методов измерения кривых плавления основан на анализе кривых, полученных методом дифференциальной сканирующей калориметрии. Обратимся снова к рисунку 2.
Пусть удельная теплота плавления липида равна Qm, а количество липодов в образце составляет m кмолей. Общее количество энергии, поглощенной образцом в интервале температур плавления T1-T2 равно очевидно площади под кривой C=f(T), т. е.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 |
