
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Ретикулярные и эластические волокна при исследовании легких инфицированных животных обнаруживали как в интерстиции, так и в зонах ателектазов и воспалительных инфильтратов во все периоды патологического процесса.
Величина объемной плотности ретикулярных волокон была больше в 2,6 раза в 1 сутки инфекционного процесса по сравнению с величиной аналогичного показателя в группе контроля. Максимального значения величина исследуемого показателя достигала на 6 сутки, то есть в 2,4 раза больше по сравнению с таковым в 1 сутки эксперимента. К 14 суткам инфекционного процесса наблюдали снижение величины показателя объемной плотности ретикулярных волокон в 2 раза по сравнению с 6 сутками эксперимента (см. табл. 2).
При исследовании эластических волокон легких инфицированных мышей, обнаружили, что показатель их объемной плотности был выше в 4,6 раза по сравнению с величиной аналогичного показателя в группе контроля. Максимальную величину объемной плотности эластических волокон наблюдали на 10 сутки, увеличение по сравнению с величиной аналогичного показателя в 1 сутки патологического процесса составило более чем в 3 раза (см. табл. 2).
Как следует из представленных данных, суммарный объем ретикулярных и эластических волокон на 3 сутки патологического процесса составил более 36% легких, тогда как коллагеновых волокон – около 7% (см. рис. 1). К 10 суткам процент «молодой» соединительной ткани – 58%, объем же коллагеновых волокон составил более 12% (см. табл. 2).
Таблица 2
Результаты морфометрического исследования содержания волокнистой соединительной ткани в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
Коллагеновые волокна (Vv), % | 1 | 0,91±0,58c | 4,6±1,29ac |
3 | 6,6±0,77ac | ||
6 | 10,4±1,08abc | ||
10 | 12,6±0,86abc | ||
14 | 16,2±1,24abc | ||
Эластические волокна (Vv), % | 1 | 1,6±0,45c | 7,4±0,71ac |
3 | 16,3±1,25abc | ||
6 | 17,6±1,01ac | ||
10 | 23,5±1,57abc | ||
14 | 22,4±1,16ac | ||
Ретикулярные волокна (Vv), % | 1 | 6,1±1,41c | 15,6±2,49ac |
3 | 20,1±2,76ac | ||
6 | 37,7±2,97abc | ||
10 | 34,3±3,36ac | ||
14 | 19,5±1,18abc |
Примечания:
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования;
«с» - достоверность отличия величин рассматриваемых параметров между разными типами волокон по периодам исследования.
При оценке соотношения величин показателей объемной плотности «молодой» и «зрелой» волокнистой соединительной ткани наблюдали постепенное уменьшение соотношения величин исследуемых показателей во все периоды инфекционного процесса (см. рис. 2).
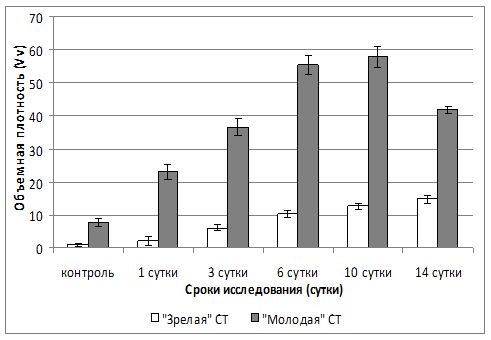
Рис 2. Результаты исследования соотношения величин объемных плотностей «зрелой» (коллагеновые волокна) и «молодой» волокнистой соединительной ткани (эластические и ретикулярные волокна) в легких мышей, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 в динамике заболевания.
Так, у интактных животных соотношение коллагеновых волокон к ретикулярным и эластическим составляло 7,85. К 1 суткам эксперимента наблюдали увеличение величины соотношения изучаемых показателей в 1,4 раза. В последующем, с 1 по 14 сутки величина соотношения «молодой» соединительной ткани к «зрелой» уменьшилась почти в 4 раза.
Результаты изучения различных типов коллагена легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05
Составной частью внеклеточного матрикса соединительной ткани являются фибриллярные структуры, которые в большей степени представлены коллагеновыми белками – коллагенами I, III, IV типов, создающими каркас всех легочных структурных образований (, 2010; Suki B., Bates J. H., 2008).
При гриппе A/H5N1 A/goose/Krasnoozerskoye/627/05 в легких экспериментальных животных уже на ранних сроках инфекционного процесса в интерстиции легких наблюдали накопление всех исследуемых типов коллагена с преобладанием коллагена III типа. На 1-3 сутки после инфицирования коллаген I и III типа обнаруживали преимущественно в межальвеолярных перегородках, периваскулярно и перибронхиально. Начиная с 6 суток инфекционного процесса, данные типы коллагена регистрировали также в зонах инфильтрации и ателектазов.
При ИГХ исследовании в легких экспериментальных животных наблюдали утолщение базальных мембран за счет увеличения в них содержания коллагена IV типа во все периоды инфекционного процесса. Также отмечали его увеличение вокруг бронхов и сосудов. Коллаген VI типа преобладал в межальвеолярных перегородках, а также в инфильтратах и зонах ателектазов, начиная с 3 суток инфекционного процесса.
Показатель объемной плотности коллагена I типа в 1 сутки после инфицирования был больше почти в 2 раза относительно аналогичного показателя в группе контроля. В последующие периоды инфекционного процесса наблюдали постепенное увеличение показателя объемной плотности коллагена I типа, с максимальным его значением на 10 сутки, что в 1,6 раза больше, чем в 1 сутки заболевания. К 14 суткам отмечали стабилизацию процесса синтеза коллагена I типа (см. табл. 3).
Величина объемной плотности коллагена III типа была выше в 1 сутки инфекционного процесса в 2,3 раза относительно аналогичного показателя в группе контроля, достигая максимальной величины на 6 сутки после инфицирования, что в 1,5 раза больше, чем в 1 сутки заболевания. В последующие сроки (10 и 14 сутки) величина объемной плотности коллагена III типа постепенно снижалась, достигая величины таковой в 1 сутки инфекционного процесса (см. табл. 3).
В легких инфицированных животных величина показателя объемной плотности коллагена IV типа была больше к 1 суткам заболевания в 1,5 раза по сравнению с величиной аналогичного показателя у мышей контрольной группы. На 3 и 6 сутки после инфицирования наблюдали постепенное увеличение исследуемого показателя, его величина была максимальной на 6 сутки после инфицирования, увеличиваясь в 1,3 раза по сравнению с его величиной в 1 сутки эксперимента. На 10 и 14 сутки наблюдали снижение величины объемной плотности коллагена IV типа (см. табл. 3).
При исследовании объемной плотности коллагена VI типа в легких мышей, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 было обнаружено, что величина его объемной плотности была больше почти в 1,5 раза по сравнению с величиной аналогичного показателя в группе контроля, с последующей стабилизацией процесса вплоть до 10 суток, когда произошло увеличение величины исследуемого показателя в 1,4 раза по сравнению с его величиной в 1 сутки инфекционного процесса. На 14 сутки заболевания величина объемной плотности VI типа коллагена достоверно не отличалась относительно таковой в предыдущий период заболевания (см. табл. 3).
Таблица 3
Результаты исследования объемной плотности коллагенов различных типов в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
Коллаген I (Vv), % | 1 | 6,2±0,92 | 12,1±1,41a |
3 | 13,3±1,12a | ||
6 | 16,1±1,34ab | ||
10 | 19,5±1,76ab | ||
14 | 17,9±1,21a | ||
Коллаген III (Vv), % | 1 | 6,2±0,74 | 14,3±1,32a |
3 | 16,45±1,40a | ||
6 | 20,6±1,35ab | ||
10 | 15,5±0,92ab | ||
14 | 12,7±0,83ab | ||
Коллаген IV (Vv), % | 1 | 8,4±0,45 | 12,4±1,16a |
3 | 12,7±0,91a | ||
6 | 16,5±1,10ab | ||
10 | 16,2±1,25a | ||
14 | 14,8±0,90a | ||
Коллаген VI (Vv), % | 1 | 5,1±0,84 | 7,4±1,22a |
3 | 8,8±1,35a | ||
6 | 7,8±0,94a | ||
10 | 10,2±1,30ab | ||
14 | 12,2±1,72a |
Примечания:
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования.
Результаты изучения структурно-функциональной активности фибробластов легких мышей линии C57Bl/6 в динамике инфекционного процесса
При микроскопическом исследовании легких инфицированных мышей увеличение количества фибробластов наблюдали периваскулярно и перибронхиально, начиная с 3 суток после инфицирования – в инфильтратах, состоящих из лимфоцитов, макрофагов и в утолщенных межальвеолярных перегородках.
Согласно результатам ИГХ исследования легких мышей, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05, в 1 сутки заболевания величина численной плотности фибробластов по сравнению с таковой у интактных животных была большей почти в 6 раз (см. табл. 4). В последующем величина этого показателя продолжала увеличиваться, достигая максимального значения на 10 сутки после инфицирования, что в 1,7 раза больше по сравнению с величиной того же показателя в 1 сутки заболевания. На 14 сутки инфекционного процесса наблюдали снижение величины численной плотности фибробластов в 1,3 раза относительно таковой в предыдущий период заболевания (см. табл. 4).
Ранее на модели с использованием беспородных мышей, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 в дозе 10 МЛД50, было показано, что вирус способен реплицироваться в альвеолоцитах, макрофагах легких и эндотелиоцитах легочных артерий ( и др., 2008). В данном эксперименте методом ИФ определяли антиген вируса A/H5N1 (Inf A), меченного флюорохромом FITC в фибробластах. Была выявлена его экспрессия в фибробластах легких инфицированных мышей уже в 1 сутки инфекционного процесса. В период с 1 по 3 сутки доля инфицированных фибробластов составляла около 25% всех фибробластов легких инфицированных мышей. Затем происходило резкое снижение величины численной плотности Inf A+фибробластов до 7% от всех фибробластов легких вплоть до 14 суток (см. табл. 4). Вероятно, это происходило в связи с гибелью части инфицированных фибробластов путем апоптоза (Daidoji T. et al., 2008; Hu Y. et al., 2012), а их относительное количество резко уменьшалось в связи с активными процессами пролиферации фибробластов (см. табл. 4), возможно, той их части, которая не была инфицирована.
Таблица 4
Результат исследования численной плотности фибробластов, экспрессирующих исследуемые маркеры в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
Фибробласты (Nai) | 1 | 1,4±0,10 | 8,2±0,86a |
3 | 14,3±0,74ab | ||
6 | 13,6±0,65a | ||
10 | 14,3±0,72a | ||
14 | 11,1±0,47ab | ||
Фибробласты Inf A+ (Nai) | 1 | - | 2,1±0,34 |
3 | 3,2±0,37 | ||
6 | 0,8±0,18b | ||
10 | 0,8±0,18 | ||
14 | 0,7±0,21 | ||
Фибробласты PCNA+ (Nai)nai) %астовская активность"авнению с предыдущим сроком исследованияспрессирующих ия. о срока исследования. последующей стабилиз | 1 | 0,9±0,21 | 2,5±0,27a |
3 | 3,7±0,29ab | ||
6 | 3,8±0,61a | ||
10 | 4,7±0,89a | ||
14 | 1,3±0,33b | ||
«Фибропластическая активность» фибробластов | 1 | 0,7 | 0,26 |
3 | 0,43 | ||
6 | 0,76 | ||
10 | 0,88 | ||
14 | 1,33 |
Примечания:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
