
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования.
Известно, что активность ММР-2 регулируется TIMP-2 (Lakatos G, et al., 2012), активность ММР-8, -9, -10, регулируется TIMP-1 (Huang L. W. et al., 2000; Clark R. T. et al., 2011). Следует принимать во внимание, что TIMP-1 является ингибитором нескольких типов ММРs, а TIMP-2 – ингибитор преимущественно MMP-2. Кроме того, при изучении экспрессии матриксных металлопротеиназ, ММР-2 из всех представленных MMPs в наибольшем количестве экспрессируется фибробластами. Поэтому для выявления характера соотношения экспрессии данных маркеров нами было предпринято исследование соотношения численных плотностей макрофагов и фибробластов, экспрессирующих ММР-2 и TIMP-2 (см. рис. 4).
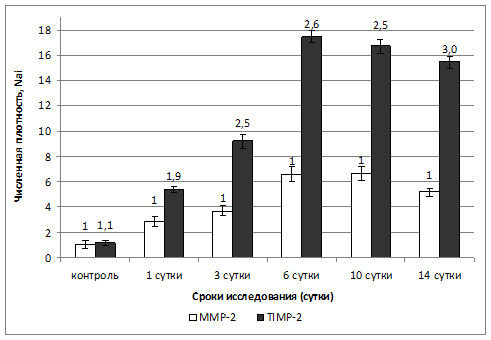
Рис 4. Результаты исследования соотношения общей численной плотности макрофагов и фибробластов, экспрессирующих MMP-2 и общей численной плотности макрофагов и фибробластов, экспрессирующих TIMP-2 в легких мышей, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 в динамике заболевания.
Как следует из представленных данных (см. рис. 4), соотношение клеток, экспрессирующих TIMP-2/MMP-2 (макрофагов и фибробластов) в легких интактных мышей равно 1,1. У инфицированных мышей исследуемое соотношение в 1 сутки эксперимента составляет 1,9, с последующим постепенным увеличением на каждом сроке инфекционного процесса. Наибольшей величины соотношение TIMP-2/MMP-2 у инфицированных мышей достигает к 14 суткам и составляет почти 3 единицы. Согласно полученным данным, преобладание численной плотности клеток, экспрессирующих TIMP-2 над численной плотностью ММР-2+клеток свидетельствуют об активации механизмов снижения у инфицированных мышей процессов внеклеточной деструкции коллагена I, IV и VI типа.
Полученные данные свидетельствуют о том, что в исследуемых экспериментальных условиях регуляция процессов синтеза и деградации структур внеклеточного матрикса реализуется фибробластами и макрофагами – «фибропластической активностью» фибробластов и протеолитической активностью MMPs, ограничивающихся, в свою очередь, избыточностью TIMP. «Дисбаланс» этих процессов в рассматриваемой ситуации, видимо, одна из причин раннего фиброза легких согласно данным корреляционного анализа (см. табл. 9).
Таблица 9
Результаты корреляционного анализа численных плотностей фибробластов и фибробластов, экспрессирующих MMPs, TIMP-2, а также фибробластов, экспрессирующих MMPs, TIMP-2 и «фибропластической активности» фибробластов в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05
Объекты корреляционного исследования | Коэффициент корреляции |
Фибробласты – MMP-2, -9, -10+фибробласты | 0,862 |
«Фибропластическая активность» – MMP-2, -9, -10+фибробласты | 0,462 |
Фибробласты – TIMP-2+фибробласты | 0,835 |
«Фибропластическая активность» –TIMP-2+фибробласты | 0,499 |
Результаты изучения клеточного состава и численной плотности клеток, экспрессирующих профибротические факторы роста и их рецепторы (FGF, FGFR, EGF, EGFR)
Ведущая патогенетическая роль в развитии фиброза принадлежит цитокинам, включая факторы роста (GF – growth factor), которые регулируют воспалительный ответ на повреждение клеток, в частности его репаративную фазу (фиброгенез) ( и др., 1993; , , 1990).
При гистологическом исследовании образцов легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05, на 1-3 сутки наблюдали увеличение численных плотностей макрофагов и фибробластов, экспрессирующих FGF, EGF и их рецепторы преимущественно в межальвеолярных перегородках, а также периваскулярно и перибронхиально. На 6-14 сутки эксперимента «положительно» окрашенные макрофаги и фибробласты появлялись, помимо указанных областей, в инфильтратах и зонах ателектазов.
При ИГХ исследовании образцов легких мышей линии C57Bl/6 инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 в 1 сутки заболевания величина численной плотности макрофагов, экспрессирующих FGF была выше по сравнению с таковой у интактных мышей более чем в 2 раза (см. табл. 10). В последующем наблюдали постепенное увеличение численной плотности FGF+макрофагов с ее максимумом на 10 сутки инфекционного процесса – увеличение исследуемого маркера почти в 4 раза относительно 1 суток эксперимента. К 14 суткам наблюдали снижение величины численной плотности макрофагов, экспрессирующих FGF в 1,3 раза по сравнению с предыдущим сроком исследования (см. табл. 10).
Величина численной плотности макрофагов, экспрессирующих FGFR к 1 суткам после инфицирования была больше величины данного параметра у интактных животных в 2,7 раза. В дальнейшем наблюдали постепенное увеличение численной плотности клеток, экспрессирующих этот маркер. К 10 суткам после инфицирования величина численной плотности FGFR+макрофагов была максимальной – увеличение в 2,2 раза по сравнению с величиной аналогичного параметра в 1 сутки исследования. В дальнейшем наблюдали тенденцию к снижению величины численной плотности макрофагов, экспрессирующих FGFR (см. табл. 10).
У мышей линии С57Bl/6 в 1 сутки эксперимента величина численной плотности EGF+макрофагов была больше величины аналогичного параметра в группе контроля в 1,8 раза. Наибольшей величина численной плотности макрофагов, экспрессирующих EGF была на 6 сутки эксперимента, превышая величину численной плотности исследуемого маркера более чем в 2 раза по сравнению с таковой в 1 сутки инфекционного процесса (см. табл. 10). На 10 и 14 сутки наблюдали снижение количества EGF+макрофагов, к 14 суткам в 1,4 раза по сравнению с величиной аналогичного показателя в 6 сутки заболевания.
Численная плотность макрофагов, экспрессирующих EGFR у инфицированных мышей в 1 сутки эксперимента была больше, чем у интактных мышей в 1,8 раза. С 1 по 10 сутки после инфицирования в легких мышей регистрировали увеличение численной плотности EGFR+макрофагов в 3,4 раза. Величина исследуемого показателя к 14 суткам эксперимента по сравнению с величиной численной плотности EGFR+макрофагов на 10 сутки эксперимента не изменялась (см. табл. 10).
Таблица 10
Результаты исследования численной плотности макрофагов, экспрессирующих исследуемые параметры в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
FGF (Nai) | 1 | 0,7±0,15 | 1,5±0,21a |
3 | 3,6±0,27ab | ||
6 | 5,5±0,32ab | ||
10 | 5,9±0,29a | ||
14 | 4,6±0,33ab | ||
FGFR (Nai) | 1 | 0,8±0,19 | 2,2±0,24a |
3 | 2,8±0,27ab | ||
6 | 4,8±0,35ab | ||
10 | 4,9±0,25a | ||
14 | 4,5±0,34a | ||
EGF (Nai) | 1 | 1,1±0,22 | 2,1±0,33a |
3 | 2,9±0,25ab | ||
6 | 4,4±0,42ab | ||
10 | 3,7±0,31a | ||
14 | 3,2±0,27a | ||
EGRF (Nai) | 1 | 0,8±0,16 | 1,5±0,23a |
3 | 2,3±0,38ab | ||
6 | 4,3±0,34ab | ||
10 | 5,2±0,27ab | ||
14 | 4,9±0,36a |
Примечания:
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования.
При гистологическом исследовании образцов легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 в 1 сутки после инфицирования величина численной плотности фибробластов, экспрессирующих FGF была больше по сравнению с таковой в контроле в 2 раза. Максимальной, величина численной плотности FGF+фибробластов была на 10 сутки эксперимента – в 2,4 раза по сравнению с величиной аналогичного показателя в 1 сутки заболевания (см. табл. 11).
Величина численной плотности фибробластов, экспрессирующих FGFR в 1 сутки исследования было больше в 2,8 раза величины аналогичного показателя у мышей в группе контроля. Количество фибробластов, экспрессирующих FGFR постепенно увеличивалось, достигая наибольшего значения на 14 сутки эксперимента. Таким образом, величина численной плотности FGFR+фибробластов увеличилась в 2,2 раза по сравнению с величиной аналогичного показателя в 1 сутки после инфицирования (см. табл. 11).
Количество фибробластов, экспрессирующих EGF в легких инфицированных мышей было больше величины аналогичного параметра у интактных мышей в 2,2 раза. В последующие сроки исследования наблюдали увеличение численной плотности EGF+фибробластов с наибольшим значением на 6 сутки эксперимента – в 1,9 раза больше по сравнению с таковой в 1 сутки инфекционного процесса (см. табл. 11). На 10 и 14 сутки эксперимента наблюдали стабилизацию величины численной плотности фибробластов, экспрессирующих EGF.
Величина численной плотности фибробластов, экспрессирующих EGFR в 1 сутки эксперимента была большей, чем в группе контроля в 1,6 раза. С 1 по 10 сутки после инфицирования в легких мышей регистрировали увеличение численной плотности EGFR+фибробластов в 3,2 раза. К 14 суткам наблюдали тенденцию к снижению исследуемого показателя (см. табл. 11).
Таблица 11
Результаты исследования численной плотности фибробластов, экспрессирующих исследуемые параметры в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
FGF (Nai) | 1 | 1,2±0,12 | 2,5±0,27a |
3 | 4,2±0,31ab | ||
6 | 5,7±0,37ab | ||
10 | 5,8±0,42a | ||
14 | 5,2±0,35a | ||
FGFR (Nai) | 1 | 0,9±0,24 | 2,7±0,28a |
3 | 3,9±0,24ab | ||
6 | 4,9±0,18ab | ||
10 | 5,5±0,33ab | ||
14 | 5,9±0,25a | ||
EGF (Nai) | 1 | 1,4±0,19 | 3,0±0,13a |
3 | 3,6±0,22ab | ||
6 | 5,5±0,27ab | ||
10 | 4,9±0,36ab | ||
14 | 5,3±0,34a | ||
EGRF (Nai) | 1 | 1,0±0,14 | 1,7±0,24a |
3 | 2,8±0,28ab | ||
6 | 4,7±0,35ab | ||
10 | 5,3±0,34a | ||
14 | 4,8±0,41a |
Примечания:
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования.
При оценке корреляционных взаимоотношений между численной плотностью фибробластов, экспрессирующих FGF, EGF и их рецепторы и численной плотностью всех фибробластов легких мышей линии C57Bl/6 наблюдали прямую высокую степень корреляционной взаимосвязи. Согласно данным корреляционного анализа между величинами показателей численной плотности фибробластов, экспрессирующих исследуемые факторы роста и их рецепторы и «фибропластической активностью» фибробластов наблюдали прямую среднюю степень взаимосвязи (см. табл. 12). Полученные данные корреляционного анализа свидетельствуют о том, что увеличение количества фибробластов и макрофагов, экспрессирующих профибротичекие факторы роста и их рецепторы в большей степени обусловливают увеличение количества фибробластов в легких инфицированных мышей, нежели на их «фибропластическую активность», значительно влияя, таким образом на объем соединительной ткани.
Таблица 12
Результаты корреляционного анализа исследуемых параметров в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05
Объекты корреляционного исследования | Коэффициент корреляции |
Фибробласты – FGF+фибробласты | 0,902 |
Фибробласты – EGF+фибробласты | 0,834 |
Фибробласты – FGFR+фибробласты | 0,846 |
Фибробласты – EGFR+фибробласты | 0,787 |
«Фибропластическая активность» – FGF+фибробласты | 0,515 |
«Фибропластическая активность» – EGF+фибробласты | 0,558 |
«Фибропластическая активность» – FGFR+фибробласты | 0,618 |
«Фибропластическая активность» – EGFR+фибробласты | 0,664 |
Коллагеновые волокна (Vv) – FGF+клетки и EGF+клетки (Nai) | 0,911 |
Коллагеновые волокна (Vv) – FGFR+клетки и EGFR+клетки (Nai) | 0,973 |
Таким образом, из полученных данных следует, что повышенная экспрессия профибротических факторов роста и их рецепторов в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05, влияющих на процессы роста, пролиферации и хемотаксиса фибробластов способствуют увеличению объема синтезируемых фибробластами компонентов внеклеточного матрикса, то есть, развитию фиброза.
Результаты исследования экспрессии провоспалительных цитокинов TNF-α и IL-6 фибробластами легких после инфицирования мышей линии C57Bl/6 вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05
При ИГХ исследовании легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 обнаружили, что интерстициальные фибробласты, а также фибробласты в составе инфильтратов демонстрировали положительную реакцию, при окрашивании на маркеры TNF-α и IL-6. Кроме фибробластов, данные маркеры также экспрессировали макрофаги, альвеолоциты и эндотелиоциты сосудов легких.
В легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 величина численной плотности фибробластов, экспрессирующих TNF-α была большей, чем у мышей в контрольной группе в 1,3 раза. Наибольшей величины исследуемый показатель достиг на 6 сутки – количество TNF-α+фибробластов увеличилось почти в 3 раза по сравнению с величиной аналогичного параметра в 1 сутки эксперимента. В последующие сроки эксперимента регистрировали снижение величины численной плотности фибробластов, экспрессирующих TNF-α в 1,3 раза на 14 сутки по сравнению с его величиной на 6 сутки эксперимента.
Количество IL-6+фибробластов в легких инфицированных мышей в 1 сутки исследования было большим в 1,7 раза по сравнению с величиной исследуемого параметра у интактных мышей. С 1 по 6 сутки наблюдали увеличение численной плотности фибробластов, экспрессирующих IL-6 в 3,3 раза, с последующим уменьшением их количества к 14 суткам в 1,45 раза (см. табл. 13).
Таблица 13
Результаты исследования численной плотности фибробластов, экспрессирующих TNF-α и IL-6 в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 (M±m)
Объект и параметры исследования | Сутки после инфицирования | Мыши линии C57Bl/6 | |
Интактные | Инфицированные | ||
TNF-α (Nai) | 1 | 1,2±0,12 | 1,5±0,19ab |
3 | 2,0±0,22ab | ||
6 | 3,4±0,21ab | ||
10 | 2,8±0,23ab | ||
14 | 2,6±0,31a | ||
IL-6 (Nai) | 1 | 0,5±0,06 | 0,9±0,18ab |
3 | 1,6±0,16ab | ||
6 | 3,0±0,31ab | ||
10 | 2,2±0,26ab | ||
14 | 2,0±0,33a |
Примечания:
«а» - достоверность отличия средних величин рассматриваемых параметров инфицированных мышей по сравнению с аналогичными показателями у мышей в контрольной группе;
«b» - достоверность отличия средних величин рассматриваемых параметров с предыдущим периодом исследования
При анализе корреляционных взаимоотношений между количеством фибробластов, экспрессирующих TNF-α, IL-6 и общим количеством фибробластов, обнаружили положительную высокую степень корреляции. Средняя корреляционная связь между количеством PCNA+фибробластов и количеством фибробластов, экспрессирующих исследуемые провоспалительные цитокины, очевидно обусловлена их слабой степенью дифференцированности (см. табл. 14). Между величинами численной плотности TNF-α+фибробластов, IL-6+фибробластов и объемной плотности коллагеновых волокон, наблюдали высокую и среднюю положительные корреляционные связи (см. табл. 14).
Таблица 14
Результаты корреляционного анализа численных плотностей фибробластов, экспрессирующих исследуемые провоспалительные цитокины в легких мышей линии C57Bl/6, инфицированных вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05
Объекты корреляционного исследования | Коэффициент корреляции |
Фибробласты (Nai) – TNF-α+фибробласты (Nai) | 0,794 |
Фибробласты (Nai) – IL-6+фибробласты (Nai) | 0,816 |
PCNA+фибробласты (Nai) – TNF-α+фибробласты (Nai) | 0,593 |
PCNA+фибробласты (Nai) – IL-6+фибробласты (Nai) | 0,623 |
«Фибропластическая активность» – TNF-α+фибробласты (Nai) | 0,512 |
«Фибропластическая активность» – IL-6+фибробласты (Nai) | 0,458 |
Коллагеновые волокна (Vv) – TNF-α+фибробласты (Nai) | 0,855 |
Коллагеновые волокна (Vv) – IL-6+фибробласты (Nai) | 0,623 |
TNF-α – провоспалительный цитокин, обладающий способностью стимулировать не только пролиферацию фибробластов, но и синтез ими коллагена (, 1998). Об этом же свидетельствуют данные корреляционного анализа (см. табл. 14), где показан более высокий коэффициент корреляции TNF-α, чем в случае IL-6.
ВЫВОДЫ
1. Инфицирование мышей линии С57Вl/6 вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05, наряду с другими клетками приводит к инфицированию фибробластов и последующему нарастанию масштаба деструктивных изменений в легких не только вследствие прямого цитопатического воздействия вирусов, но и его косвенно опосредованных эффектов, детерминирующих процессы локальной гипоксии и ишемии вследствие тромбоэмболических, геморрагических осложнений, нарушения проницаемости стенки гемических сосудов, интерстициального и внутриальвеолярного отеков.
2. Инфицирование фибробластов вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 является триггерным фактором, «запускающим» процессы пролиферации фибробластов на ранних этапах после инфицирования животного и увеличивающим долю дедифференцированных фибробластов в легких, у которых по мере продолжительности периода после инфицирования усиливается «фибропластическая активность» как структурно-функциональное проявление их дифференцированности.
а) Инфицирование фибробластов легких вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 является триггерным фактором процессов синтеза фибробластами структур внеклеточного матрикса и их последующей трансформации, приводящей к раннему фиброзированию легких, поскольку масштабы его и пролиферации фибробластов увеличивались вопреки существенному уменьшению численности инфицированных вирусом гриппа фибробластов.
б) Сохраняющаяся персистенция вируса во многих типах клеток, в частности фибробластах и макрофагах, препятствует восстановлению структурно-функционального гомеостаза.
3. Инфицирование мышей линии C57Bl/6 вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 сопряжено с активацией синтеза и избыточным отложением в легких коллагенов I, III, IV, VI типов, свидетельствующих о том, что процесс их фиброзирования носит не только заместительный (репаративно-постнекротический), но и индуцированный состоянием гипоксии характер, как следствие нарушения процессов газообмена в интерстиции, через стенки и базальные мембраны гемических сосудов
4. В условиях острой вирусной инфекции, индуцированной вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 у мышей линии С57В1/6 процессы синтеза и протеолиза структур внеклеточного матрикса в легких детерминированы численностью и «фибропластической активностью» фибробластов, а также численностью фибробластов и макрофагов, экспрессирующих матриксные металлопротеиназы (ММРs): MMP-2, MMP-9, MMP-10.
а) Дисгармоничные отношения между процессами синтеза и деградации структур внеклеточного матрикса, могут быть, в частности, обусловлены избыточной экспрессией ТIМР-2, ингибирующего чрезмерную активность ММРs как механизм гомеостазирования внеклеточного матрикса, а также уменьшения риска повреждения его конституитивных структур в рассматриваемой экспериментальной модели, что может быть одним из вероятных факторов, детерминирующих фиброзирование легких.
б) Высокое содержание коллагена III типа, в том числе в составе ретикулярных и эластических волокон, в легких мышей после их инфицирования вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 и резкое снижение его содержания на 10 и 14 сутки после инфицирования на фоне постепенного существенно менее масштабного увеличения содержания в легких коллагена I типа, дает основание для предположения о том, что в рассматриваемой экспериментальной модели «молодая» соединительная ткань более предпочтительно и интенсивно деградируется ММРs, чем «зрелая», а часть ее «расходуется» на формирование «зрелых» волокон соединительной ткани.
5. Инфицирование фибробластов вирусами гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 сопряжено с активацией ими экспрессии ростовых факторов и их рецепторов (FGF, FGFR, EGF, EGFR), стимулирующих пролиферацию фибробластов, что в итоге приводит к увеличению общего объема коллагена и иных структур внеклеточного матрикса легких.
а) Увеличение количества макрофагов в легких, экспрессирующих те же факторы роста и их рецепторы, что и фибробласты, может стимулировать хемоаттракцию фибробластов в локусы деструкции в тех зонах органа, где недостаточно содержание конституитивных фибробластов на момент повреждения для их эффективной репарации по типу substitucio, что возможно, является фактором риска развития фиброза легких.
6. В условиях инфицирования мышей вирусом гриппа A/H5N1 A/goose/Krasnoozerskoye/627/05 фибробласты легких способны поддерживать процесс воспаления, экспрессируя провоспалительные цитокины TNF-a и IL-6, стимулируя, таким образом, свою пролиферацию, вследствие чего, по мере их дифференцирования, возрастает риск усиления фиброзирования органов.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Особенности фибропластических процессов в легких мышей оппозитных линий CBA и C57Bl/6 при гриппе A/H5N1 / , , , // Фундаментальные аспекты компенсаторно-приспособительных процессов: Материалы 5-й Всероссийской научно-практической конференции с международным участием. – 2011, Новосибирск. – С. 11.
2. Морфофункциональные изменения эндотелиоцитов сосудов легких при гриппе А/H5N1 у млекопитающих / , , // Клинические и теоретические аспекты современной медицины: Материалы IV Международной студенческой научной конференции с участием молодых ученых. – 2012, Москва. – С. 49–50
3. Структурно-функциональные изменения легочных макрофагов и легких мышей, инфицированных вирусом гриппа А/H5N1 A/goose/krasnoozerskoye/627/05 / , , , , // Бюллетень экспериментальной биологии и медицины. – 2012. – № 2. – С. 196–199. (Из списка ВАК).
4. Аникина, А. Г. Морфофункциональная характеристика фибробластов и особенности фиброгенеза в легких мышей, инфицированных вирусом гриппа А/H5N1 A/goose/Krasnoozerskoye/627/05 / , , // Бюллетень экспериментальной биологии и медицины. – 2012. – № 9. – С. 346–349. (Из списка ВАК)
Автор благодарит сотрудников лаборатории отдела зоонозных инфекций и гриппа под руководством , ФГУН ГНЦ ВБ «Вектор» (Новосибирск) за ведение совместного эксперимента.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
