
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Цистенілтрансфераза
Н2О
![]() СО2
СО2
![]() 4'-фосфопантетеїн В6-Ф 4'-фосфопантотеніл цистеїн
4'-фосфопантетеїн В6-Ф 4'-фосфопантотеніл цистеїн
![]() Декарбоксилаза
Декарбоксилаза
![]() АТФ
АТФ
ФФн Аденілтрансфераза
![]()
![]()
 дефосфо-КоА коензим А
дефосфо-КоА коензим А
Кіназа
АТФ АДФ
Рисунок 11 – Схема біосинтезу коензиму А
Нормальна концентрація та поширення у тканинах. Вміст вітаміну В5 у плазмі крові становить 15 – 45 мг/100 мл. У невеликій кількості він наявний у всіх тканинах організму. Найбільші концентрації відмічені у печінці, серці, нирках.
Екскреція. Продукти катаболізму пантотенової кислоти невідомі. За умов нормального харчування із сечею виділяється від 2,5 до 5 мг/добу вітаміну В5, із потом екскретується 3 – 4 мг/100 мл, секретується з молоком 200 – 300 мг/100 мл.
Вітамін В5 значно поширений у тканинах рослин та тварин.
Джерела:
· продукти тваринного походження: печінка, нирки, яєчний жовток, знежирене молоко, сир, курятина, риба;
· продукти рослинного походження: хлібні злаки, боби, хліб із висівками, солодка картопля;
· дріжджі;
· більшість овочів і фруктів бідні на вітамін В5;
· близько 57 % вітаміну руйнується під час виготовлення борошна та 33 % при кулінарній обробці м'яса;
· більше ніж 3,4 мг на добу синтезується мікрофлорою кишечнику, за відсутності дисбактеріозу.
Добова потреба у пантотеновій кислоті невідома через її значне поширення у природі.
Рекомендоване добове споживання на 2500 кал:
· дорослі: 5 – 12 мг;
· новонароджені: 1 –2 мг;
· діти: 4 – 5 мг;
· вагітні жінки 15 – 20 мг.
Потреба у вітаміні підвищується при гострих захворюваннях, опіках, серйозних пораненнях, лікуванні антибіотиками, під час видужання після хвороби, у період вагітності та лактації. Для забезпечення достатньої кількості вітаміну у дітей, що інтенсивно ростуть, у період формування мозку та інших внутрішніх органів, молоко матері повинно вмішувати до 5 мг пантотенової кислоти на 1 л. Потреба у вітаміні В5 збільшується при стресових станах, великих фізичних навантаженнях, заняттях спортом.
Біологічна роль
Біологічна роль пантотенової кислоти пов'язана з її входжен-ням до структури коферменту А (коферменту ацилювання), який бере участь у реакціях активації та перенесення ацетильного (СН3С=О) залишку та радикалів інших кислот. Внасладок утво-рення ацил-КоА карбонова кислота активується, складаються вигідні термодинамічні умови для її використання у ендерго-нічних процесах. Як компонент HS-КоА вітамін В5 необхіден для перебігу декількох фундаментальних біохімічних процесів.
1. Формування «активного» ацетату (ацетил-КоА). Коен-зим А легко взаємодіє з ацетатом із утворенням метаболічно активного Ацетил-КоА:
О
║
СН3 - С ~ S-КоА.
Ацетил-КоА містить у своїй структурі високоенергетичний сульфатний зв'язок, еквівалентний макроергічному фосфату АТФ. Близько 80 ферментів залежать від пантотенової кислоти. У формі активного ацетату вітамін бере участь у:
· циклі трикарбонових кислот – у реакції з оксалоацетатом утворює цитрат;
· реакціях ацетилювання;
· синтезі нейтральних жирів, ацетилхоліну, фосфоліпідів, холестеролу, кетонових тіл;
· біосинтезі та елонгації жирних кислот;
· β-окисненні жирних кислот;
· окиснювальному декарбоксилюванні пірувату та α-кетоглу-тарату;
· утворенні стероїдних гормонів кори надниркових залоз;
· активації амінокислот із розгалуженими ланцюгами – ізолейцину, лейцину, валіну;
· біосинтезі гіпурової кислоти.
2. Формування «активного» сукцинату (сукциніл-КоА). Сукциніл-КоА – продукт окиснювального декарбоксилювання α-кетоглутарату у циклі лимонної кислоти, який бере участь у нижче наведених метаболічних процесах:
· Синтезі гему. Сукциніл-КоА взаємодіє з гліцином і утворює δ-АЛК - ключовий метаболіт синтезу гему.
· Розщепленні кетонових тіл у позапечинкових тканинах.
У людини ізольований дефіцит пантотенової кислоти виникає рідко через її значне поширення у харчових продуктах і біосинтез мікрофлорою ШКТ. Недостатність вітаміну В5 проявляється у вигляді:
· дерматитів, депігментації волосся, алопеції;
· виснаження, зупинення росту, анемії;
· ураження слизових оболонок;
· дистрофічних змін залоз внутрішньої секреції, зокрема надниркових залоз, серця, нирок, ШКТ;
· порушень нервової системи – невритів, паралічів;
· печіння у стопах є характерним проявом дефіциту вітаміну.
Основні причини гіповітамінозу В5:
· захворювання ШКТ, при яких відбувається порушення природної мікрофлори кишечнику або всмоктування вітаміну;
· довготривале нераціональне харчування, лікування антибіотиками;
· часто є супутником інших гіповітамінозів.
Пантотенову кислоту використовують для лікування дерматитів, захворювань печінки, серця, у парфумерії.
4.5. Вітамін В6
(піридоксин, адермін, антидерматитний)
Хімічна структура та властивості
· За структурою вітамін В6 є 2-метил-3-оксі-4,5-діоксиметил-піримідином. Відповідно до Міжнародної номенклатури тер-міном «вітамін В6» позначають три похідні 3-оксипіридину, які мають однакову вітамінну активність: піридоксин (піридоксол), піридоксаль, піридоксамін.
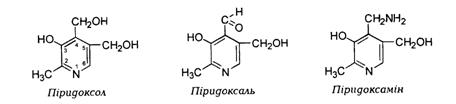
· Вітамін В6 добре розчинний у воді та етанолі.
· Водні розчини вітаміну стійкі до кислот і лугів, але чутливі до дії ультрафіолетового світла при нейтральному рН.
· Вітамін стійкий до температури, але втрачається під час консервування.
Біологічно активні форми
Коферментні властивості притаманні лише фосфорильова-ним формам піридоксалю та піридоксаміну – ПАЛФ (піридоксальфосфат) і ПАМФ (піридоксамінфосфат). У природі вони зустрічаються в основному в комплексі з білками.
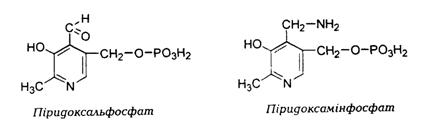
Фосфорилювання піридоксалю та піридоксаміну відбуваєть-ся у печінці, мозку та інших тканинах за участі АТФ, Zn++ і специфічних кіназ.
Утворення піридоксальфосфату:
Дегідрогеназа Кіназа, Zn++
![]()
![]()

![]() Піридоксин Піридоксаль Піридоксаль-Ф
Піридоксин Піридоксаль Піридоксаль-Ф
Фосфопротеїн Фосфопротеїн-Н2 АТФ АДФ+ Фн
Вітамін В6 синтезується багатьма мікроорганізмами і росли-нами. В організмі людини він не утворюється, але у невеликій кількості синтезуватися мікроорганізмами кишечника.
Всмоктування. Вітамін В6 легко абсорбується у тонкому ки-шечнику простою дифузією і розноситься кров’ю до тканин. Частина – депонується у зв'язаному з білком вигляді, переважно у печінці, більшість перетворюється у коферментні форми.
Екскреція. Піридоксаль і піридоксамін у невеликій кількості (0,5 - 0,7 мг/добу) виводяться з сечею. Основний біологічно неактивний метаболіт, який екскретується - 4-піридоксинова кислота (3,0 мг/добу).
Вітамін значно поширений у тканинах рослин і тварин. У рослинних продуктах переважає піридоксин, а у тваринних – піридоксаль та піридоксамін. У багатьох продуктах тваринного походження піридоксин знаходиться у комплексі з білками, але у ШКТ під дією ензимів він легко вивільняється і всмоктується.
Джерела:
· продукти рослинного походження: дріжджі, хліб, морква, червоний перець, картопля, горох, квасоля, крупи, капуста. У наших умовах основними джерелами вітаміну В6 є хліб та картопля;
· продукти тваринного походження: м'ясо, печінка, нирки, риба, жовток яйця.
Добова потреба у піридоксині для людини точно не встановлена, оскільки частково потреби у ньому задовольня-ються за рахунок синтезу мікрофлорою кишечнику. Розрахована добова потреба становить близько 2,0 мг/добу. Вважається, що вона залежить від надходження білків з їжею, оскільки кофермент бере участь у багатьох реакція обміну амінокислот.
Добова потреба:
· дорослі: 2,0 мг;
· новонароджені: 0,3 – 0,4 мг;
· жінки у другій половині
вагітності: 2,5 мг.
Потреба значно підвищується при довготривалому лікуванні туберкульозу ізоніазидом, який є антивітаміном вітаміну В6.
Біологічна роль
Піридоксальфосфат діє як кофермент, в основному у реакціях амінокислотного обміну.
· Трансамінування. ПАЛФ і ПАМФ, що взаємоперетворю-ються, є коферментами амінотрансфераз у зворотних реакціях трансамінування амінокислот.
· Декарбоксилювання. Кофермент (ПАЛФ) декарбоксилаз у реакціях незворотного декарбоксилювання амінокислот із утворенням відповідних біогенних амінів:
o Тирозин → Тирамін + СО2.
o Гістидин → Гістамін + СО2.
o Глутамінова кислота → ГАМК + СО2.
· Кофермент деаміназ (дегідраз), які каталізують реакції нео-киснювального дезамінування гідроксіамінокислот серину та треоніну.
· Кофермент кінуренінази, що перетворює 3-ОН-кінуренін на 3-ОН-антралову кислоту, з якої утворюється нікотинова кис-лота. Тому при нестачі В6 синтез ніацину з триптофану припиняється. У крові підвищується рівень кінуреніну та 3-ОН-кінуреніну, які у позапечінкових тканинах перетворю-ються на ксантуринову кислоту. Остання виділяється з сечею, що є надійним критерієм діагностики дефіциту В6.
· Транссульфурування. Бере участь у перенесенні сульфгід-рильних (-SH) груп, наприклад, у реакції:
Гомоцистеїн + Серин → Гомосерин + Цистеїн.
· Кофермент дисульфурази, яка каталізує неокиснювальне дезамінування цистеїну, при якому виділяється H2S.
· Бере участь у взаємоперетвореннях гліцину та серину серин-гідрокси-метилтрансферазою.
· Біосинтезі арахідонової кислоти з ліноленової.
· Синтез сфінгомієліну. ПАЛФ – кофермент у активації серину, останній необхідний для синтезу сфінгомієліну.
· Кофермент рацемаз амінокислот:
D-Глутамінова кислота → L-Глутамінова кислота
D-Аланін → L-Аланін.
· Внутрішньомітохондріальний синтез жирних кислот – кофермент елонгації жирних кислот у мітохондріях.
· Необхідний для «активного транспорту» амінокислот через клітинну мембрану та їх абсорбції у кишечнику.
· Структурний компонент фосфорилази м'язів: чотири молекули піридоксальфосфату разом із однією молекулою ензиму утворюють тетрамерну структуру.
· Сприяє транспорту іонів К+ через мембрану у клітини.
· Діє як кофермент аміноацетонсинтетази в утворенні аміноацетону з ацетил-КоА та гліцину.
· Синтез КоА-SH з пантотенової кислоти. При гіповітамінозі В6 вміст коензиму А у печінці значно знижується.
· Синтез порфіринів. Піридоксальфосфат є коферментом δ-амінолевулінатсинтази – ключового ферменту синтезу ге-му. Недостатність вітаміну викликає розвиток анемії.
У людини гіповітаміноз В6 зустрічається рідко і не описано якоїсь специфічної хвороби його недостатності. Дефіцит піри-доксину краще вивчений на тваринах.
Клінічні та біохімічні прояви недостатності вітаміну В6:
у щурів спостерігається акродинія – специфічний дерматит, який вражає шкіру лапок, хвіст, ніс, вуха, дегенеративні зміни у ЦНС.
У людини:
· епілептиформні напади у немовлят, яких вигодовують незбалансованими за вітаміном В6 штучними сумішами. Зниження активності ПАЛФ-залежної глутаматдекарбок-силази призводить до зменшення утворення тормозного медіатора γ-аміномасляної кислоти у мозку, підвищення збудливості нервової системи, що є причиною судом.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
