
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
· Вітамін А і β-каротин, як потужні антиоксиданти викорис-товують для профілактики та лікування онкологічних захворювань. Вони перешкоджають повторній появі пухлини після операцій.
· Антиоксидантні властивості β-каротину лежать в основі використання його у профілактиці захворювань серця та судин.
· Каротиноїди лютеїн і зеаксантин застосовують для профі-лактики хвороб очей, зокрема розвитку катаракти.
· Ретиноїди та їх синтетичні аналоги використовують у сучасних косметичних засобах для підтримки і відновлення епітелію шкіри.
· Вітамін Е та рибофлавін (вітамін В2) підсилюють дію вітаміну А, тобто є його фізіологічними синергістами.
3.2. Вітаміни групи D
Синоніми: кальциферол, "аксерофтол", антирахітний вітамін, «вітамін сонячного світла».
Хімічна структура та властивості
Група вітаміну D об'єднує декілька хімічних сполук, які роз-різняються за хімічною будовою та біологічною активністю, їх неактивні попередники «провітаміни» містяться у природних продуктах:
· вітамін D2 – ергокальциферол (рослинного походження), його провітамін – ергостерол;
· вітамін D3 – холекальциферол (тваринного походження), його провітамін – 7-дегідрохолестерол (синтезується у шкірі);
· вітамін D4 – 22,23-дигідроергокальциферол;
· вітамін D5 – етилхолекальциферол (ситокальциферол); виділений з пшеничної олії;
· вітамін D6 – 22-дигідроетилкальциферол (сигма-кальци-ферол):
Вітамін D2 – ергокальциферол та вітамін D3 – холекальциферол є найбільш поширеними природними формами вітаміну D, тоді як інші представники цієї групи є його модифікованими формами.
За хімічною будовою попередники вітаміну D – це одноатомні ненасичені циклічні спирти, в основі їх структури лежить ядро циклопентанпергідрофенантрену. Провітаміни D містять:
· -ОН групу біля С3;
· два кон’югованих подвійних зв’язки між С5-С6 та С7-С8;
· вуглеводневий ланцюг біля С17.
Вітаміни D2 та D3 – кристали без кольору і запаху, стійкі до впливу високих температур, розчинні у жирах і органічних розчинниках, не розчинні у воді.
Вітамін D2 утворюється у рослинах із ергостеролу. Він знайдений у опромінених дріжджах.
В організмі людини вітамін D3 утворюється у дермальному шарі шкіри з попередника – провітаміну D3 (7-дегідрохолестеро-лу). Сам провітамін D утворюється з холестеролу у печінці та кишечнику, транспортується в шкіру, де під впливом коротко - хвильового ультрафіолетового опромінення (довжина хвилі 290-315 нм) трансформується у вітамін D3. Внаслідок дії сонячного світла та температури тіла відбувається фотохімічна реакція розкриття В кільця стероїдного ядра і термоізомеризація попередника. Механізм фотохімічної активації провітамінів:
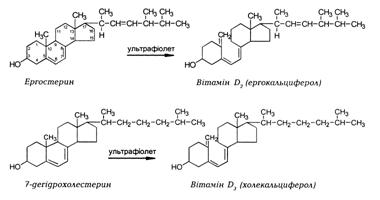
Всмоктування. Вітамін D3 легко засвоюється у кишечнику і після всмоктування у складі хіломікронів транспортується через лімфу у кров до печінки. Ергостерол погано засвоюється з їжі. 7-Дегідрохолестерол синтезується у клітинах кишечнику з холестеролу, переноситься у шкіру для біотрансформації на вітамін D3. Як більшість жиророзчинних сполук, жовчні кислоти сприяють всмоктуванню вітаміну D у кишечнику.
Утворення біологічно активних форм. При фізіологічних умовах кльцифероли неактивні. Вітамін D виконує свої функції у формі біологічно активних метаболітів, які утворюються з нього у печінці та нирках:
· 1,25-діоксихолекальциферол [1,25(ОН)2D3];
· 24,25-діоксихолекальциферол [24,25(ОН)2D3];
· 25-оксихолекальциферол є попередником вищеназваних метаболітів (транспортною формою кальциферолів).
Біологічно активною формою вітаміну D є гормон кальцитріол [1,25(ОН)2D3], який синтезується у печінці та нирках. В організмі здорової дорослої людини утворюється близько 0,3 – 1,0 мкг гормону на добу.
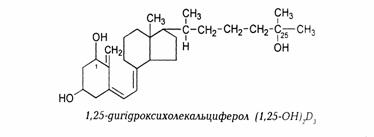
(а) Перша стадія активації вітаміну D - синтез 25-ОН-D3 (кальцидіолу) відбувається переважно у печінці (рис.5):
· Вітаміни D2 та D3 у складі специфічного вітамін-D-зв'язувального протеїну транспортуються у печінку. Для нормального метаболізму вітаміну D у гепатоцитах необхіден вітамін Е.
· У ендоплазматичному ретикулумі гепатоцитів вітамін D гідроксилюється у положенні 25 під дією мікросомальної НАДФН-залежної цитохром Р-450 редуктази (25-гідроксилази). Коферменти та кофактори, що викорис - товуються у активації:
o Mg++, НАДФН, молекулярний О2.
· Реакція 25 – гідроксилювання відбувається дуже швидко, що призводить до підвищення рівня 25-ОН-D3 у сироватці крові та інгібування активністі 25-гідроксилази за принци-пом негативного зворотного зв'язку.

7-дегідрохолестерол Сонячне світло Вітамін D3
![]()
 (провітамін D3) УФ (Холекальциферол)
(провітамін D3) УФ (Холекальциферол)
О2 У печінці
![]() НАДН2
НАДН2
Mg++ 25-Гідроксилаза
25-ОН-D3
(25-ОН-Холекальциферол)
![]() У мітохондріях нирок
У мітохондріях нирок
НАДФН2, О2, Mg++
1,25(ОН)2D3![]()
![]() (Кальцитріол) 1α-Гідроксилаза
(Кальцитріол) 1α-Гідроксилаза
1,25-ді-ОН холекальциферол
Рисунок 5 – Синтез кальцитріолу
(б) Друга стадія активації вітаміну D - синтез 1,25-ди-ОН-D3 (кальцитріолу) у нирках:
- 25-ОН-D3 транспортується у нирки специфічним D-зв’я-зувальним протеїном, гідроксилюється у 1-му положенні під дією мікросомальної монооксигенази (1α-гідрокси-лази) у проксимальних канальцях нирок. У процесі беруть також участь:
- НАДФН2, О2, Mg++; додатково необхідні: фероредоксинредуктаза, феродоксин, цитохром Р450.
(в) Регуляція синтезу кальцитріолу:
o кальцитріол інгібує активність 1α-гідроксилази за принци-пом негативного зворотного зв'язку, стимулює утворення 24,25-ді-ОН-D3 та експресію специфічного ензиму – D–24–гідроксилази. Останній каталізує перетворення каль-цитріолу на неактивну водорозчинну кальцитроєву кислоту, яка екскретується з жовчу.
o Паратгормон активує синтез кальцитріолу у нирках.
o Рівнями фосфатів та кальцію у сироватці крові: гіпокаль-цемія підвищує активність 1α-гідроксилази за наявності паратгормону.
o 1α-Гідроксилаза активується естрогенами, андрогенами, кальцитоніном, пролактином, соматотропіном, глюкокор-тикостероїди, навпаки, виявляють інгібуючий ефект.
Поширення у тканинах. Рівень 25-ОН-D3 у крові відобра-жає як утворення вітаміну D у шкірі, так і його надходження з їжею, у зв'язку з чим може використовуватися як маркер статусу вітаміну. З крові вітамін D проникає у печінку, кістки, скелетні м'язи, нирки, надниркові залози, міокард, жирову тканину. основна форма накопичення вітаміну D в організмі та його. Особливо довго вітамін зберігається у адипоцитах і м'язах, створюючи тканинні депо.
Екскреція. Вітамін екскретується з жовчю у кишечник, з якого частково реабсорбується.
Вітамін D2 значно поширений у рослинах, але погано всмоктується у кишечнику, тому не може розглядатися як важливе джерело вітаміну D.
Джерела:
· продукти тваринного походження є кращими джерелами вітаміну D3: риб'ячий жир, вершкове масло, жовток яйця, печінка риб та тварин;
· продукти рослинного походження: найбільш багаті на вітамін D2 рослинні олії (соняшникова, оливкова), петрушка, кропива, дріжджі, гриби;
· молоко та молочні продукти не можуть задовольнити потре-бу організму у вітаміні D через низький вміст його у цих продуктах та високу концентрацію фосфору, що перешкод-жає засвоєнню кальциферолів.
Ендогенний синтез на 80 % задовольняє потреби організму у вітаміні (за умов достатньої інсоляції) і залежить від:
o довжини хвилі світла (найбільш ефективний середній спектр хвиль, який ми отримуємо вранці та на заході сонця);
o вихідної пігментації шкіри (чим темніше шкіра, тим менше вітаміну D виробляється);
o віку – синтез вітаміну зменшується з віком;
o рівеня забруднення атмосфери – промислові викиди та пил не пропускають ультрафіолетових променів.
Добова потреба у вітаміні D для дорослих людей становить 2,5 – 10,0 мкг/добу. Вважається, що короткочасне (10 – 30 хви-лин) сонячне опромінення обличчя і відкритих рук еквівалентне добовій потребі у вітаміні.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
