
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
o У зворотній реакції перетворення гліцину на серин «1-С» атом переміщується на H4-фолат, формується N5, N10- метилен-FH4. У зв'язку з тим, що гліцин здатен синтезуватися з глюкози, остання може бути джерелом одновуглецевого фрагменту (1).
o Перенесення одновуглецевого фрагменту від гліцину на H4-фолат призводить до утворення СО2, NH4+(2).
· У результаті катаболізму гістидину синтезується форміміно-глутамат. У реакції останнього з H4-фолатом вивільняється NH4+, утворюється глутамат і N5, N10-метеніл-FH4 (4).
· N10- форміл-FH4 може синтезуватися з триптофану (5).
Похідні ТГФК є донорами одновуглецевих фрагментів у реакціях синтезу багатьох речовин (табл. 4):
· N10- форміл-FH4, N5,N10-метеніл-FH4 є донорами атомів карбону для утворення С2 та С8 атомів пуринового ядра.
· N5, N10- метилен-FH4 переносить «1-С» групу на дУМФ з утворенням дТМФ. У цій реакції метиленова група відновлюється до метильної, а тетрагідрофолева кислота окиснюється до дегідрофолевої. Регенерація H4-фолату відбуається шляхом відновлення під дією НАДФН2-залежного ензиму дегідрофолатредуктази.
· N5, N10- метилен-FH4 є також донором вуглецю під час перетворення Глі на Сер.
· Переміщення метильної групи з N5-метил-FH4 на вітамін В12 призводить до утворення метилкобаламіну, який, у свою чергу, є донором СН3 – у реакціі ресинтезу метіоніну з гомоцистеїну. У подальшому метильна група активної форми метіоніну - S-аденозилметіоніну бере участь у синтезі таких біологічно активних сполук, як катехоламіни, креатин, фосфоліпіди, мелатонін та ін.
Недостатність фолієвої кислоти важко викликати навіть у експериментальних тварин через її значне поширення у їжі та достатній синтез мікрофлорою кишечнику. Штучний дефіцит вітаміну у тварин можливий при використанні спеціального раціону без фолієвої кислоти при одночасному довготривалому вживанні сульфаніламідних препаратів та антибіотиків широкого спектра дії, які інгібують ріст кишкової мікрофлори і, таким чином, пригнічують власний синтез фолієвої кислоти; використанні антагоністів вітаміну Вс.
Клінічні прояви дефіциту фолієвої кислоти:
· порушення кровотворення і розвиток специфічної макроци-тарної анемії, яка за клінічними ознаками подібна до перніціозної анемії, викликаної дефіцитом вітаміну В12, але без нервових порушень;
· зниження кількості еритроцитів і гемоглобіну. Поява у пери-ферійній крові молодих клітин – мегалобластів із низьким вмістом ДНК, як наслідок, порушення синтезу ДНК у еритропоетичних клітинах кісткового мозку;
· зменшення кількісті лейкоцитів (лейкопенія);
· розвиток хейлозу, специфічного глоситу (сухий червоний язик), кон'юктивіту;
· наслідком гастриту та ентериту, які розвиваються на фоні недостатності вітаміну, спостерігається ахлоргідрія, діарея, стеаторея;
· затримка росту, імунодифіцитні стани, зниження швидкості загоєння ран, хронічні інфекції.
Біохімічні зміни при авітамінозі Вс:
· порушення включення одновуглецевих фрагментів у різні сполуки;
· неможливість синтезу пуринових нуклеотидів і піримідинової азотистої основи тиміну. Тимідилові нуклео-тиди перестають включатися у структуру ДНК і частково замінюються на уридилові, як наслідок, синтезується аномальна ДНК;
· зниження швидкості синтезу ДНК призводить до порушення клітинного циклу, у першу чергу в швидко проліферуючих клітинах – епітеліальних, гемопоетичних;
· порушення синтезу серину, валіну, ресинтезу метіоніну з гомоцистеїну, накопичення останнього спричиняє недостат-ність метіоніну;
· уповільнення катаболізму гістидину призводить до акумулювання проміжного метаболіту – форміміноглута-мату. Визначення цього деривату в сечі використовується як діагностичний тест для диференціювання дефіциту вітаміну Вс від В12 недостатності.
Основні причини гіповітамінозу фолієвої кислоти:
· тривала недостатність харчування (відсутність у їжі свіжих сирих продуктів) та теплова обробка їжі впродовж тривалого часу. Часто дефіцит вітаміну розвивається при неповноцінному харчуванні, наприклад у «бутербродників», хронічних алкоголіків;
· мегалобластична анемія вагітних може виникати у жінок (особливо у ІІІ триместрі вагітності) через підвищену потребу у вітаміні;
· дефіцит вітаміну в матері призводить до передчасних пологів і є причиною розвитку мегалобластичної анемії у новонароджених. У подальшому такі діти відстають у фізичному та розумовому розвитку;
· «сімейний тип» мегалобластичної анемії, яка розвивається через уроджений дефект активності редуктази;
· підвищення потреби у фолієвій кислоті виникає при низькому вмісті білка у раціоні, хронічних захворюваннях печінки та нирок, гемодіалізі, лейкемії, гемолітичній анемії, хронічних інфекціях, псоріазі, у курців.
· гіповітаміноз вітаміну В12, при якому клітини легко втрачають фолієву кислоту і не здатні її використовувати через неможливість деметилювання фолацину та збереження його у інтрацелюлярному просторі;
· макроцитарна анемія може розвинутися при хронічній діареї, стеатореї, целіакії, спру після резекції кишечнику, через неадекватну реабсорбцію вітаміну;
· анемія, викликана дефіцитом вітаміну Вс, може виникнути у людей похилого віку;
· довготривале лікування такими препаратами, як метотрексат, протисудомні та протимікробні засоби, оральні контрацеп-тиви та ін.
· Аміноптерин: за хімічною структурою є 4-амінофолатом. Він має максимальну інгібуючу дію, блокує утворення коферментної форми вітаміну В9.
· Аметоптерин, або метотрексат: структурно це 4-аміно-10- метилфолат.
Обидва антивітаміни блокують синтез нуклеїнових кислот шляхом інгібування відновлення фолевої кислоти до тетрагідропохідних фолієвої кислоти.
Вітамін використовують для лікування мегалобластичної та післярезекційної анемії, спру, анемії, яка викликана дією радіації та ліків, отруєння етанолом, стимулювання еритропоезу, посилення репаративних процесів.
4.7. Вітамін В12
(кобаламін, антианемічний, зовнішній фактор Касла)
Хімічна структура та властивості:
· за хімічною будовою вітамін належить до класу кориноїдів;
· молекула його складається із двох частин: кобальтовмісної порфіриноподібної (хромофорної) групи – «корзинового ядра» та нуклеотидної структури;
· нуклеотидна частина представлена 5,6-диметил-бензимід-азолрибонуклеотидом;
· В12 – єдиний металовмісний вітамін, який у центрі ядра хромофорної структури містить Со;
· кобальт з’єднаний із атомами азоту відновлених пірольних кілець і з атомом азоту 5,6-диметилбензимідазолу. Останній поєднаний із рибозою та залишком фосфорної кислоти. Вся структура має назву «кобалaмін»;
· пірольні кільця містять різні радикали – метильні, ацетамідні, пропіонамідні;
· були також отримані похідні вітаміну В12, що вміщують ОН - групу, Сl-, NO2, SO42-, які відповідно мають назви: гідроксокобаламін, хлоркобаламін, нітритокобаламін, сульфокобаламін;
· з природних джерел також були виділені менш біологічно активні аналоги вітаміну В12, які замість 5,6-диметил-бензимідазолу вміщували 5-оксибензимідазол, або аденін, 2-метиладенін, гіпоксантин, метилгіпоксантин.
Біологічно активні форми
Коферментні форми вітаміну В12 (кориноїдні коферменти) синтезуються в печінці та нирках у серії послідовних реакцій за участі специфічних ензимів, АТФ, НАД, ФАД, глутатіону. Утворюються два типи коферментів – метилкобаламін та 5'-дезоксіаденозилкобаламін, які містять відповідно метильну групу або 5'-дезоксіаденозин (рис. 12, 13).
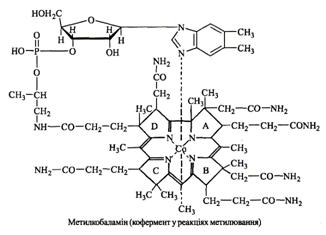
Рисунок 12 – Коферментна форма вітаміну В12 – метилкобаламін
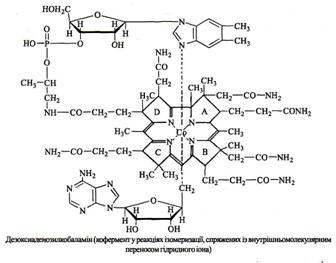
Рисунок 13 – Коферментні форми вітаміну В12 - 5'-дезоксиаденозил-
кобаламін
Біосинтез
Вітамін В12 синтезується виключно мікроорганізми і не утворюється у клітинах рослин чи тварин.
Метаболізм
Всмоктування. Вітамін В12 всмоктується у тонкій кишці за наявності соляної кислоти, іонів Са++ та «внутрішнього фактора Касла» (ФК) – гастромукопротеїну, який містить у своєму складі гексози, гексозамін, сіалові кислоти.
Механізм всмоктування охоплює (рис. 14):
· утворення комплексу вітаміну В12 із фактором Касла (ФК) для перенесення останнього через клітини слизової оболонки тонкої кишки;
· зв'язування комплексу В12-фактор Касла зі специфічними рецепторами на мембрані ентероцитів;
· перенесення комплексу через мембрану, відділення вітаміну В12 від внутрішнього фактора за участі іонів Са++ та вивільнювального фактора.

 ФК ФК
ФК ФК![]() Са2+
Са2+
![]()
![]()
![]()
![]()
![]()
![]()
 В12 → В12 → В12 В12
В12 → В12 → В12 В12
![]()
![]()
![]()
![]() Кров
Кров
В12
Транскобаламін
Рисунок 14 – Схема всмоктування вітаміну В12 у тонкому
кишечнику
· гідроліз фактора Касла у клітинах кишечнику;
· всмоктування кобаламіну у портальний кровотік і зв'язуван-ня його з транспортними білками α2- і β-глобулінів крові – транскобаламіном І та транскобаламіном ІІ.
· транспортування вітаміну В12 у клітини печінки для депонування;
· від 0,5 до 10,0 мг вітаміну В12 всмоктується завдяки вище-названому механізму. Близько 1 – 3 % кобаламіну абсорбу-ється шляхом простої дифузії.
Поширення у тканинах. Основне місце депонування вітаміну В12 – печінка, у якій створюється запас вітаміну, що може задовольнити фізіологічні потреби організму впродовж 3 – 5 років після зникнення внутрішнього фактора у шлунку. Значна кількість його накопичується також у нирках і селезінці. Загальний вміст вітаміну в організмі людини становить близько 2 – 5 мг. Метаболізм кобаламіну відбувається дуже повільно, тому при поступовому зниженні його надходження в організм ознаки гіповітамінозу розвиваються тільки через 5 – 6 років.
Нормальна концентрація у плазмі крові: 200 – 750 пг/мл.
Виділення. Зазвичай вітамін не виділяється із сечею, але при парентеральному введенні добова екскреція дорівнює близько 0,3 мг. Вітамін В12 екскретується з жовчю, але більша частина його знову всмоктується, тобто для нього характерна ентерогепатична циркуляція.
Вітамін наявний виключно у продуктах тваринного походження і не міститься у рослинній їжі, оскільки синтезується тільки мікроорганізмами у кишечнику тварин.
Деяка кількість вітаміну міститься у бобових культурах, де його продукують мікроорганізми.
Джерела:
· продукти тваринного походження, багаті на вітамін В12: печінка, нирки, яєчний жовток, м'ясо, риба;
· продукти тваринного походження з помірним вмістом вітаміну В12: молоко та молочні продукти;
· у невеликій кількості вітамін синтезується мікрофлорою кишечнику, за умов надходження з їжею кобальту. Однак цей синтез не може повністю забезпечити потреби організму через погану абсорбцію вітаміну з ШКТ.
Добова потреба у вітаміні В12 для дорослих становить близько 3 мкг/добу.
Рекомендоване споживання:
· дорослі: 3,0 мкг;
· новонароджені: 0,3 мкг;
· діти: 1 – 2 мкг;
· жінки у період вагітності
та лактації: 4,0 мкг;
- при перніціозній анемії
парентеральне введення 0,5 – 1,0 мг.
Біологічна роль
Коферментні форми вітаміну В12 беруть участь у двох типах реакцій: ізомеризації та трансметилювання. Встановлені тільки 2 метаболічні реакції, у яких безпосередньо беруть участь коферментні форми вітаміну В12.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
