


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Некоторые исследователи отмечают, что синезеленые водоросли могут использовать углекислоту в таких концентрациях, при которых другие водоросли теряют способность к размножению, поэтому они нередко явно доминируют в водоемах с высокими значениями рН и низким содерджаниемуглекислоты [136]. Однако, как показано в работе [137], подщелачиване среды при поглощении углекислоты синезелеными водророслями начинает ингибировать другие виды фитопланктона. Концентрация углекислоты может оказывать влияние на синезеленыые, изменяя величину рН, что приводит к ряду неблагоприятных процессов, в частности развитию болезнетворных бактерий, перемещению цианоагов, увеличению активнсти ферментов цианобактерий, а также к геохимическим последствиям в виде изменения растворимости металлов и трансформации фосфатных форм в воде [138].
Считается, что величины рН в воде водохранилищ, изменяющиеся как во времени в течении вегетационного сезона, так и по акватории водохранилища контролируют изменения видового состава доминирующих синезеленых водорослей в периоды «цветения», поскольку для разных родов характерны различные оптимумы рН (для Anabaena 7–8, для Aphanizomenon – 8,1–8,45, для Microcystis – 9–10 [132, 139].
К одному из наиболее сложных вопросов проблемы биогенного питания фитопланктона следует отнести влияние ионов металлов на развитие синезеленых водорослей. Сложность оценок этого влияния состоит в активном участии ионов металлов в реакциях с метаболитами водорослей и другими компонентами биотического круговорота, что приводит к комплексообразованию, и, как следствие, резко изменяются формы соединений металлов и их токсичность. Это существенно затрудняет оценку последствий действия конкретных концентраций металлов на отдельные виды фитопланктона. Сложно также анализировать данные, полученные разными авторами, т. к. условия проведения опытов существенно различались, что при изучении реакции тестовых организмов на действие токсикантов может приводить к различным интерпретациям полученных результатов [140]. Вообще говоря, важность этого вопроса трудно переоценить. т. к. неоспоримо, что сукцессии водорослей под действием тяжелых металлов может привести к существенной перестройке экосистемы водоема.
В этом обзоре необходимо выделить глубокие экспериментальные, полевые и теоретические работы группы московских гидробиологов под руководством , проведенные с целью изучить реакцию фитопланктона на концентрации биогенов и их соотношений [141, 142, 143, 144]. Коротко сформулировать выводы этих исследований можно следующим образом:
- численность видов определяется абсолютными концентрациями биогенов, а относительное доминирование отдельных видов контролируеьия как раз соотношением концентраций оснвных биогенов;
- потребность конкретного вида в биогенных веществах определяет оптимальные для каждого такого вида отношения биогенов в окружающей водной среде;
- лимитирующий элемент способен полностью подавить размножение клеток отдельного вида, если он полностью потреблен в процессе развития клеток фитопланктона;
- в случае, когда потребности в каждом биогенном элементе определены для каждого вида сообщества, можно рассчитать диапазоны лимитирующих концентраций биогенных элементов;
- клетки фитопланктона обнаруживают различную потребность в зависимости от стадии развития фитопланктона (процесс «потребление-рост») и различаются для разных видов.
1.4 Биотические факторы первичного продуцирования
1.4.1 Фактор конкуренции
При анализе пространственно-временных закономерностей и количественном описании конкретных фитопланктонных сообществ необходимо учитывать одновременное воздействие различных биотических и абиотических факторов на процессы роста и смертности водорослей. Между обитателями биоценозов, в том числе фитопланктона, имеются многочисленные и разнообразные связи. Определяющими факторами в этих связях являются межвидовая конкуренция, физиологическое состояние водорослей и пищевые отношения, которые могут быть регуляторами численности фитопланктона на фоне воздействия абиотических факторов его развития.
В таблице 1 представлены типы биотических взаимодействий, из которых наиболее изучены конкуренция, хищничество, паразитизм. Рассмотрим некоторые аспекты этих взаимодействий, имеющие значения для фитопланктона (частично они рассмотрены в уже имеющемся обзоре).
Таблица 1 – Классификация взаимодействий между популяциями двух видов
№ | Типы взаимодействий | 1-й вид | 2-й вид | Общий характер взаимодействия |
1 | Комменсализм | + | 0 | Комменсал (1) получает пользу от объединения, а для 2-й популяции объединение безразлично |
2 | Мутуализм | + | + | Взаимодействие благоприятно для обоих видов |
3 | Нейтрализм | 0 | 0 | Ни одна из популяций не влияет на другую |
4 | Аменсализм | – | 0 | Популяция 2 подавляет популяцию 1, но сама не испытывает отрицательного воздействия (например, выделение токсинов синезелёными) |
5 | Паразитизм | + | – | Паразит (1) обычно мельче хозяина (2) |
6 | Хищничество (и поедание растений) | + | – | Хищник (1) обычно крупнее жертвы (2) |
7 | Конкуренция | – | – | Взаимное подавление обоих видов |
Примечание – плюс означает благоприятное воздействие, минус – неблагоприятное, ноль – воздействие отсутствует.
Конкуренция за лимитирующий пищевой ресурс – один из важнейших биотических факторов, определяющих структуру фитопланктона в водоеме.
В биологии считается, что два одинаковых в экологическом отношении и потребностях вида не могут сосуществовать в одном месте и рано или поздно один конкурент вытесняет другого. Это так называемый принцип исключения или принцип Гаузе.
На уровне особи внутри‑ и межвидовая конкуренция совпадают (в смешанной культуре водорослей все клетки, независимо от видовой принадлежности, конкурируют за элементы питания). Но результаты внутри‑ и межвидовой конкуренции различны.
Внутривидовая конкуренция водорослей регулирует популяцию и поддерживает ее плотность на стабильном уровне (точнее происходят колебания вокруг него) – на уровне плотности насыщения. Об интенсивности внутривидовой конкуренции можно судить по снижению скорости роста популяции (увеличение смертности, снижение рождаемости) при увеличении плотности. Однако ни одна популяция не может расти экспоненциально очень долго. Рано или поздно популяция должна столкнуться с неподходящими условиями обитания (при расселении) или с нехваткой ресурсов, необходимых для дальнейшего размножения. Экспоненциальный рост довольно быстро сменяется спадом численности или выходом ее на плато, резким или плавным. В последнем случае наблюдается S‑образная кривая роста популяции (часто наблюдается в экспериментах и при вселении видов в новые местообитания) (рисунок 7) [145].
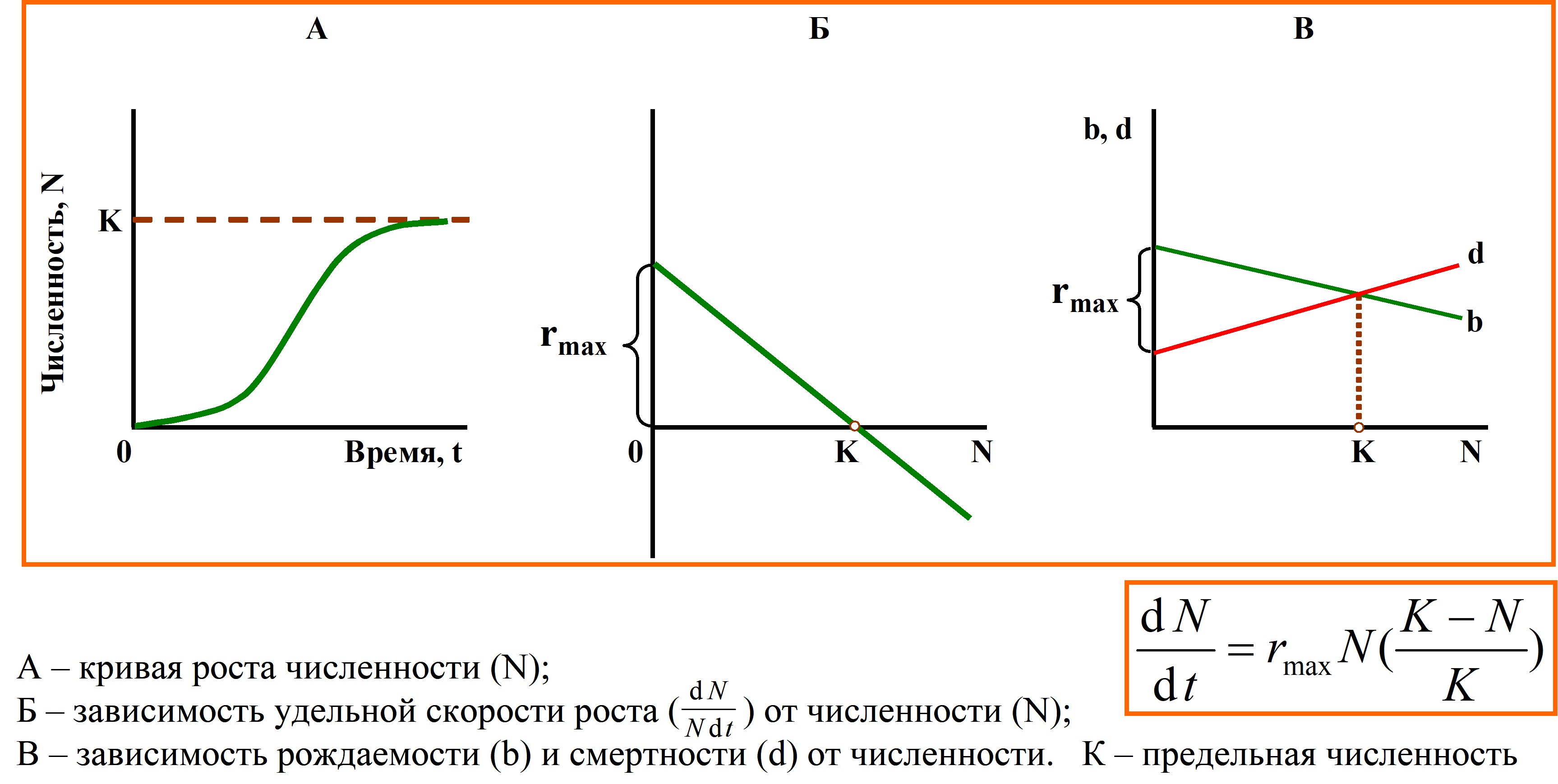
Рисунок 7 – Логистическая модель роста популяции водорослей (по: [145])
Межвидовая конкуренция водорослей не приводит к коэволюции конкурентов (что имеет место в случае взаимодействия хищника с жертвой). Выжить можно только либо избегая конкуренции (миграция в другое местообитание, переход на другой ресурс), либо развивая свою конкурентоспособность (усиливая давление на конкурента). Таким образом, результатом межвидовой конкуренции является либо расхождение конкурентов по разным экологическим нишам, либо вытеснение одного вида (конкурентное исключение) [146].
Различные популяции фитопланктона часто сосуществуют в одном и том же месте обитания, что неоднократно показывалось гидробиологическими исследованиями. Более того, многие из видов принадлежат к одной таксономической группе, поэтому характеризуются сходными реакциями на внешние воздействия, т. е. находятся в одинаковых экологических условиях. Классик современной лимнологии Г. Хатчинсон считает, что закон конкурентного исключения для фитопланктона не действует, и называет это «парадоксом фитопланктона» [147].
Регулирование численности планктонных водорослей в зависимости от особенностей метаболитных процессов изучалось в экспериментах [148]. Он пришел к выводу, что метаболиты могут считаться важнейшим фактором регулирования численности популяции. В условиях конкуренции за питание функция метаболизма – это тот механизм, который позволяет сохраниться популяции вплоть до момента естественного спада численности. Возможно, что «парадокс фитопланктона» Г. Хатчинсона объясняется именно этими причинами. Вероятно, сформировавшийся в результате эволюции механизм регулирования приводит к сложным и разнообразным пространственно-временным структурам распределения фитопланктона, существенно повышающим устойчивость планктонных сообществ к вариациям среды.
Несмотря на то, что «парадокс фитопланктона» принят большинством гидробиологов, существует немало экспериментальных доказательств существования конкурентной борьбы за продукты питания. Например, в экспериментальной работе [149] установлено, что водоросли конкурируют друг с другом за разные формы азота по‑разному, к тому же на исход конкуренции сильно влияет освещенность. Различия в освещенности и в концентрациях разных азотсодержащих веществ в зависимости от глубины и сезона (весной больше нитратов, летом – мочевины) позволяют конкурирующим видам планктонных водорослей выработать различающиеся адаптации и предпочтения и таким способом «разойтись» в экологическом пространстве и снизить конкуренцию. Благодаря этому в водоеме могут благополучно сосуществовать многие виды планктонных водорослей, несмотря на то, что всем им одинаково необходимы растворенные в воде соединения азота. Наиболее полно эти вопросы рассмотрены гидробиологами МГУ и опубликованы в цикле статей и монографий [142, 144, 150, 151, 152].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 |
